温度对马口鱼幼鱼耗氧率、窒息点及抗氧化能力的影响
来源:优秀文章 发布时间:2023-04-14 点击:
张 凯,王伟瑜,赵丽悦,熊洛梅,梅叶涵,黄可欣,刘芳玲,于 瑾,郑善坚
( 1.浙江师范大学 化学与生命科学学院,浙江 金华 321004;
2.浙江省野生动物生物技术与保护利用重点实验室,浙江 金华 321004;
3.龙游县养殖业发展中心,浙江 衢州 324000 )
鱼类作为一种水生生物,其正常生命活动均需要水的参与,温度与氧气作为水体的重要监测指标,直接影响鱼类生存、生长和繁殖,因此具有重要的研究意义。温度的高低决定着鱼体内各种酶的活性,对鱼类的新陈代谢起着决定性的作用,在适宜的温度范围内,鱼类的代谢水平与温度正相关;
当温度超出适宜范围,会影响其体内代谢水平和抗氧化水平,进而减弱鱼体清除自由基的能力,造成鱼类机体损伤[1-2]。同时,大量研究表明,低氧环境也严重抑制鱼类基础生理代谢,导致鱼类生长缓慢,机体免疫力下降,造成鱼类死亡[3-4]。
马口鱼(Opsariichthysbidens)属鲤形目鲤科马口鱼属[5-6],是亚洲分布较广的小型鲤科鱼类之一[7],该鱼在中国长江、黄河等各流域中均广泛分布,在越南、朝鲜半岛的河流中也有发现。马口鱼喜欢栖息于山间溪流中,尤其是水流较急的浅滩。由于马口鱼生性凶猛,以小鱼、昆虫为食,曾威胁淡水渔业资源,但随着水环境的污染,加之人为的过度捕捞,其自然资源数量也急剧减少。迄今为止,为保护这一鱼类资源,国内外学者进行了马口鱼生物学特征[5]、遗传特征[6]、人工驯化和培育[8-15]等相关研究,为马口鱼人工繁育技术的发展和完善提供了大量理论基础。目前,很多研究集中在马口鱼遗传育种、胚胎发育和组织结构等方面,而对幼鱼期的基础生理研究较少。为此,笔者探讨不同温度和溶解氧水平对不同规格马口鱼幼鱼耗氧率、窒息点和抗氧化指标的影响,以期为马口鱼的幼鱼期养殖生产管理提供更完善的科学依据。
1.1 试验材料
试验用马口鱼幼鱼(1000尾)由浙江渔老大农业科技有限公司提供,试验前暂养于100 L的养殖桶中,水温(24±1) ℃,每日8:00、18:00各投喂1次饲料(粗蛋白40%),暂养期间,每日换水1次,日换水量为50%。
1.2 试验方法
1.2.1 耗氧率、窒息点的测定
本试验采用静水呼吸室法,在5 L呼吸瓶(图1)中进行,试验用水为经曝气12 h[溶解氧质量浓度(8.61±0.50) mg/L]的自来水。试验前1 d停止投喂,使幼鱼肠胃完全排空。将马口鱼幼鱼根据体质量分为(4.91±0.55) g和(1.22±0.31) g 2种规格,每个呼吸瓶放入10尾马口鱼幼鱼,设20、24、28 ℃ 3个温度梯度,每个温度设3个平行组和1个空白对照组。马口鱼幼鱼放入呼吸瓶后,置于控温水槽中逐步调温至试验温度后适应2 h,随后用液体石蜡密封。

图1 试验装置结构
分别在试验前和试验开始后2 h采用虹吸法取各呼吸室上、中、下层水样于溶解氧瓶中,固定后测定其溶解氧质量浓度,并求平均值。以计算不同温度下2种规格马口鱼幼鱼的耗氧率。测量马口鱼幼鱼窒息点(试验鱼半数死亡)时的溶解氧质量浓度[16-17],认定死亡的标准为幼鱼静卧瓶底,鳃盖停止运动[18]。溶解氧质量浓度测定采用碘量法(GB 17378.4—2007)。耗氧率的计算方法参照文献[22,28]的方法:
ρ=c×V1×8×1000÷V2
式中,c为Na2SO4标准液的浓度(mol/L),8为1/2氧的摩尔质量(g/mol),V1为消耗的Na2SO4标准液的量(mL),V2为滴定时吸取的水样体积(mL),ROC为耗氧率[mg/(g·h)],ρ1、ρ2分别为初、末溶解氧质量浓度(mg/L),m为试验时鱼的质量(g),t为试验进行的时间(h)。
1.2.2 抗氧化指标的测定
分别于试验开始前[溶解氧质量浓度(8.61±0.50) mg/L]和低氧状态[溶解氧质量浓度(2.31±0.41) mg/L]时,在各呼吸瓶中随机选取3尾幼鱼进行麻醉和解剖,取其肝脏用于抗氧化指标的测定。本试验所测抗氧化指标包括总抗氧化能力、超氧化物歧化酶和丙二醛。所用试剂盒为南京建成生物工程研究所的试剂盒。
1.3 试验数据处理与分析
利用SPSS 26和Excel软件进行数据分析,通过单因素方差分析检验不同温度和规格对马口鱼幼鱼耗氧率、窒息点及抗氧化指标影响的显著性,组间差异的显著性采用LSD多重比较检验,显著水平为0.05,本试验数据均用平均值±标准差表示。
2.1 马口鱼幼鱼的耗氧率
试验结果表明:在20、24、28 ℃ 3个温度梯度下(1.22±0.31) g马口鱼幼鱼的耗氧率随温度的升高而升高,20 ℃时低至(0.185±0.057) mg/(g·h),显著低于24 ℃和28 ℃时的耗氧率(P<0.05),但24 ℃和28 ℃时的耗氧率差异不显著(P<0.05);
而(4.91±0.55) g马口鱼幼鱼的耗氧率随着温度的升高呈先升后降的趋势,24 ℃时的耗氧率(0.550±0.022) mg/(g·h)显著高于20、28 ℃温度下的耗氧率(P<0.05),且20、28 ℃时的耗氧率也存在显著差异(P<0.05);
同时也发现,除28 ℃下,(4.91±0.55) g马口鱼幼鱼耗氧率显著高于(1.22±0.31) g幼鱼(P>0.05)(表1)。将2种规格马口鱼幼鱼与本试验室测定马口鱼成鱼耗氧率(未发表)相比发现(图2),成鱼耗氧率显著低于幼鱼耗氧率(P<0.05)。
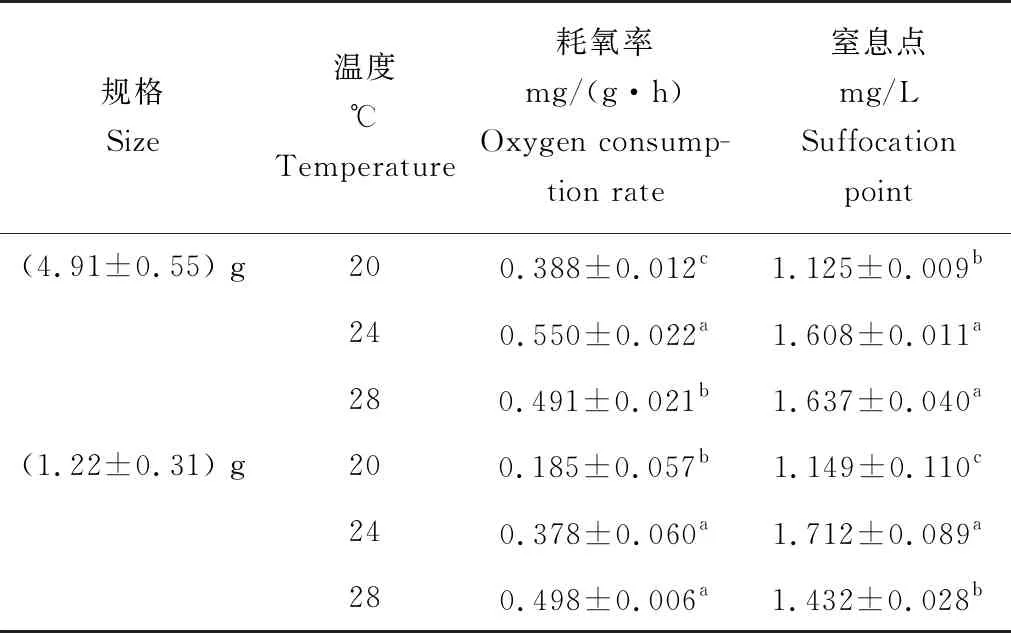
表1 不同温度下2种规格马口鱼幼鱼的耗氧率及窒息点
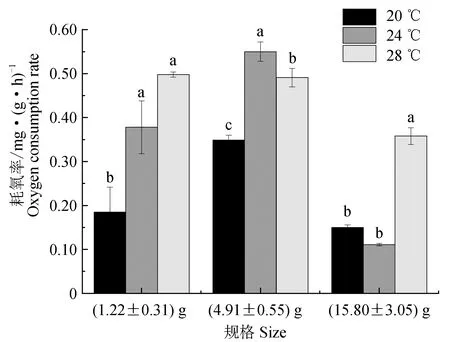
图2 不同温度下3种规格马口鱼的耗氧率
2.2 马口鱼幼鱼窒息点及行为状态
随着溶解氧质量浓度的降低,幼鱼呼吸频率明显有加快的趋势,运动频率逐渐降低,聚集在呼吸瓶底。随着溶解氧质量浓度的进一步下降,幼鱼开始出现浮头状态,呼吸频率进一步加快,部分幼鱼失去平衡,沉入瓶底,随后又快速游至水面。当呼吸瓶中溶解氧质量浓度达到窒息点时,幼鱼静卧瓶底,无呼吸,鳃和嘴张开,且部分鳃发紫,胸鳍向外伸展。测定其窒息点时的溶解氧质量浓度发现:(4.91±0.55) g马口鱼幼鱼在24和28 ℃时的窒息点差异不显著(P>0.05),20 ℃的窒息点[(1.125±0.009) mg/L]显著低于24和28 ℃的(P<0.05);
(1.22±0.31) g马口鱼幼鱼在20、24、28 ℃水温条件下的窒息点存在显著差异(P<0.05),表现为24 ℃>28 ℃>20 ℃;
2种规格马口鱼幼鱼的窒息点在20 ℃和24 ℃下差异不显著(P>0.05),28 ℃时(4.91±0.55) g幼鱼窒息点显著高于(1.22±0.31) g幼鱼(P<0.05)(表1)。在24 ℃下,将(4.91±0.55) g马口鱼幼鱼的耗氧率与北方须鳅(Barbatulabarbatulanuda)、赤眼鳟(Squaliobarbuscurriculus)、刀鲚(Coilianasus)和中华胭脂鱼(Chinesesucker)进行对比,发现马口鱼耗氧率高于相近体质量的北方须鳅、赤眼鳟和中华胭脂鱼,但窒息点显著低于刀鲚这类对低氧耐受能力差的经济鱼类(表2)。

表2 几种鱼类幼鱼窒息点的比较
2.3 低氧对马口鱼幼鱼的抗氧化影响
通过测定不同温度下马口鱼幼鱼在正常溶解氧[(8.61±0.50) mg/L]和低溶解氧[(2.31±0.41) mg/L]状态下肝脏的抗氧化指标发现,无论在正常溶解氧状态还是低溶解氧状态,(4.91±0.55) g马口鱼幼鱼在20、24、28 ℃时的超氧化物歧化酶活性随着温度的升高而升高,20 ℃时的超氧化物歧化酶活性均显著低于24、28 ℃时(P<0.05),24、28 ℃时(4.91±0.55) g幼鱼的超氧化物歧化酶活性虽有上升趋势,但差异不显著(P>0.05)(表3、表4)。20 ℃时(1.22±0.31) g幼鱼的超氧化物歧化酶活性同样显著低于24、28 ℃时(P<0.05),24、28 ℃时(1.22±0.31) g幼鱼的超氧化物歧化酶活性无显著差异(P>0.05)。
正常溶解氧下,马口鱼幼鱼20 ℃和24 ℃时丙二醛含量无显著差异,但28 ℃时(1.22±0.31) g幼鱼丙二醛含量显著高于(4.91±0.55) g幼鱼;
低氧状态下,除28 ℃下的(1.22±0.31) g幼鱼,其他幼鱼丙二醛含量高于正常溶解氧下的含量。
低氧状态下,2种规格马口鱼幼鱼在20 ℃时的总抗氧化能力无显著性差异(P>0.05),在24、28 ℃时,(4.91±0.55) g马口鱼幼鱼的总抗氧化能力约为(1.22±0.31) g马口鱼幼鱼的2倍(表3、表4)。将低氧和正常溶解氧状态下的总抗氧化能力对比发现,低氧状态下总抗氧化能力显著高于正常溶解氧(P<0.05);
比较2种规格幼鱼的总抗氧化能力发现,(4.91±0.55) g幼鱼显著高于(1.22±0.31) g幼鱼(P<0.05)。

表3 正常溶解氧[(8.61±0.50) mg/L]环境下2种规格马口鱼幼鱼的抗氧化指标

表4 低氧[(2.31±0.41) mg/L]环境下2种规格马口鱼幼鱼的抗氧化指标
3.1 水温对不同规格马口鱼耗氧率的影响
温度是影响鱼类呼吸代谢的主要因素之一[18-24]。在20 ℃和24 ℃温度下,马口鱼幼鱼的耗氧率随温度升高而显著升高,当温度达到28 ℃时,(4.91±0.55) g马口鱼幼鱼耗氧率显著降低,而(1.22±0.31) g马口鱼耗氧率有升高的趋势,但差异不显著。这说明高温对马口鱼的呼吸代谢有一定的抑制效果,与段娇阳等[25-27]的研究结果基本一致。鱼类是一种变温动物,体温随水温的变化而变化,水温的升高提高了鱼体内的酶活性,使其代谢旺盛,耗氧率也随之增加;
但当水温超出适宜范围时,其新陈代谢被抑制,耗氧率也随之降低[18]。本试验结果显示,相同温度下不同规格的马口鱼幼鱼耗氧率也存在显著差异,在20 ℃和24 ℃温度下,(4.91±0.55) g马口鱼幼鱼耗氧率显著高于(1.22±0.31) g马口鱼幼鱼,与何亚等[28-30]的研究结果存在一定差异。在已有的研究中,普遍认为鱼的耗氧率随体质量的增加而降低,主要是因为幼鱼生长迅速,基础代谢高,需要更多的营养物质来维持自身的能量消耗,相比于成鱼,幼鱼脏体比例更高,耗氧率高,随着幼鱼生长,其基础代谢降低,脏体比例逐渐减小,耗氧率也随之降低,因此,幼鱼的耗氧率相对较高[31]。经与本实验室测定的马口鱼成鱼耗氧率相比发现,成鱼耗氧率显著小于幼鱼耗氧率,但本试验中(4.91±0.55) g幼鱼耗氧率高于(1.22±0.31) g幼鱼,这可能与其生长代谢水平有关。本试验所用2种规格的马口鱼为同一批次人工繁育的幼鱼,是在人工饲养期间出现的大小分化,其表现出的对耗氧率的差异可能与其性别差异有关。唐英[32]在对马口鱼的研究中发现,同规格马口鱼雄性的生长速度高于雌性。马口鱼是典型的雌、雄异形鱼类,其成鱼阶段雄鱼与雌鱼体型存在显著差异[13,33],但幼鱼期雌、雄个体在外部形态上差异小,性染色体分化程度较低,且性别分化也易受环境影响,导致性表型与遗传基础不一致的现象[34],因此对马口鱼幼鱼期雌雄性别分化和生长性能之间的关系的研究造成了一定的阻力。同一批幼鱼在相同的养殖环境下出现不同的规格,表明其生长性能有一定的差异,进而表现出幼鱼耗氧率的差异。
3.2 水温对不同规格马口鱼窒息点的影响
鱼类窒息点是反映其代谢水平的重要依据[35]。窒息点反映了鱼类对低氧环境的耐受能力,尤其是在水产养殖、运输操作过程中具有重要的指导意义[36]。本试验结果表明,(4.91±0.55) g马口鱼幼鱼的窒息点随温度的升高而升高,这与徐林通等[27,29,37-38]在其他经济鱼类的研究中的结果相似,而(1.22±0.31) g马口鱼幼鱼的窒息点有随温度升高而升高的趋势,但28 ℃时出现了下降。在20 ℃时,2种规格的马口鱼幼鱼的窒息点较低,表明马口鱼幼鱼在低温的情况下对低氧的耐受能力强。(4.91±0.55) g马口鱼幼鱼在24 ℃和28 ℃条件下,其窒息点无显著差异,而(1.22±0.31) g马口鱼幼鱼在28 ℃下窒息点显著降低,说明28 ℃对于(1.22±0.31) g马口鱼幼鱼的呼吸代谢有显著抑制效果,由此可见,(4.91±0.55) g马口鱼幼鱼对于高温的耐受能力高于(1.22±0.31) g马口鱼幼鱼。由于本试验温度梯度设置范围较大且只有3个梯度,因此具体低温耐受范围还需进一步探究。马口鱼是一种溪流型经济鱼类,栖息在流动性较大和清澈的水域中,对水质的要求较其他鱼类幼鱼要高,因此在养殖过程中应充分做好溶解氧管理,以避免幼鱼期因缺氧导致死亡率剧增。
3.3 低氧胁迫下不同水温对马口鱼抗氧化指标的影响
鱼类抗氧化指标与温度之间具有相关性,因此被广泛用于评价鱼类新陈代谢的能力和生理健康程度[43-44]。活性氧是调控鱼类正常生命活动的重要因素,在正常的情况下会保持动态平衡,但在环境因子(温度、溶解氧质量浓度等)的胁迫下,鱼体内的活性氧平衡被打破,对鱼体造成氧化压力[45]。在低氧环境下,鱼体内的黄嘌呤脱氢酶可转化为黄嘌呤氧化酶,线粒体电子传递链载体失活,导致活性氧含量增加[46-47]。为应对活性氧的失调,鱼类形成了抗氧化系统,如超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶等一系列抗氧化酶。丙二醛是脂质过氧化作用的最终分解产物,其含量可间接反映机体的活性氧自由基和脂质的过氧化水平,从而间接反映出细胞受损伤的程度,该指标多用于水生态毒理学的研究[48-49]。总抗氧化能力是近年研究发现的用于衡量机体抗氧化系统功能状况的综合性指标,代表和反映机体抗氧化酶系统和非酶促系统对外来刺激的代偿能力以及机体自由基代谢的状态[45]。在低氧环境下[溶解氧质量浓度(2.31±0.41) mg/L],马口鱼幼鱼的超氧化物歧化酶活性随温度的升高而升高,但在28 ℃时,(1.22±0.31) g马口鱼幼鱼的超氧化物歧化酶活性有降低的趋势,表明高温对(1.22±0.31) g马口鱼幼鱼的抗氧化能力有一定的抑制效果,且(4.91±0.55) g马口鱼幼鱼的超氧化物歧化酶活性显著高于(1.22±0.31) g的马口鱼幼鱼,表明(4.91±0.55) g马口鱼幼鱼的抗氧化能力强于(1.22±0.31) g马口鱼幼鱼;
与正常溶解氧[(8.61±0.50) mg/L]相比,低氧环境下马口鱼的超氧化物歧化酶活性明显较高,表现出了马口鱼机体对于低氧环境的适应。在低氧状态20 ℃下,(1.22±0.31) g马口鱼幼鱼的丙二醛含量显著高于(4.91±0.55) g马口鱼幼鱼,表明其在低氧胁迫下,(1.22±0.31) g马口鱼幼鱼更容易受到低氧环境的损伤;
低氧情况下幼鱼的丙二醛含量显著高于正常溶解氧水平下的含量,表现出在低氧环境下,幼鱼更易受到损伤。在20 ℃和28 ℃低氧环境下(4.91±0.55) g马口鱼幼鱼总抗氧化能力和正常溶解氧环境的总抗氧化能力无显著差异;
而24 ℃时,低氧环境下的(4.91±0.55) g马口鱼幼鱼总抗氧化能力显著高于正常溶解氧,此时,低氧下的(4.91±0.55) g马口鱼幼鱼抗氧化能力更强,对环境的耐受能力也更强。将2种规格幼鱼的总抗氧化能力进行对比,发现(4.91±0.55) g马口鱼幼鱼总抗氧化能力高于(1.22±0.31) g马口鱼幼鱼,说明(4.91±0.55) g马口鱼幼鱼抗氧化系统强于(1.22±0.31) g马口鱼幼鱼。
与其他鱼类相似,马口鱼幼鱼的耗氧率和窒息点随温度升高呈先升后降趋势,不同的是马口鱼窒息点较高,且在低氧和温度胁迫下易受到损伤。因此,养殖过程中应重点关注养殖环境的适宜性,以期在较高的存活率下,创造更大的经济价值。
猜你喜欢耗氧率马口幼鱼非遗视野下湖北马口窑的保护、继承与开发流行色(2019年5期)2019-12-13北方须鳅的耗氧节律、耗氧率和窒息点的研究水产科学(2018年3期)2018-06-04——马口窑文献与当代陶艺创作研究展">来自马口窑的对话——马口窑文献与当代陶艺创作研究展美术文献(2016年3期)2016-07-05对话马口窑中华手工(2016年6期)2016-07-04马口煤矿分层开采工作面上覆采空区自燃治理技术研究中国煤炭(2016年9期)2016-06-15黄颡鱼幼鱼的赖氨酸需要量动物营养学报(2015年10期)2015-12-01黄颡鱼幼鱼对饲料中维生素C的需要量动物营养学报(2015年10期)2015-12-01不同生态因子对日本矶海绵耗氧率的影响河北渔业(2015年7期)2015-07-21不同温度条件下褐菖鲉幼鱼的耗氧率和排氨率应用海洋学学报(2014年1期)2014-11-22温度和盐度对厚壳贻贝耗氧率的影响浙江海洋大学学报(自然科学版)(2014年6期)2014-03-13推荐访问:幼鱼 抗氧化 窒息