苹果丝裂原活化蛋白激酶MdMKK9互作蛋白筛选与验证
来源:优秀文章 发布时间:2023-01-21 点击:
庄婕,杨美香,李欣欣,王彦博,孙晓红,3,张玉刚,4
(1.青岛农业大学园艺学院,山东青岛 266109;
2.青岛市城阳第四中学,山东青岛266109;
3.青岛农业大学生命科学学院;
4.山东省园艺作物基因改良工程实验室,山东青岛 266109)
丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号级联系统广泛存在于真核生物体内,是一种信号传递系统,负责将胞外多种生物或非生物胁迫的刺激信号传递至胞内,引起胞内应答[1];
在信号传递过程中,MAPK级联信号系统能够将信号整合、放大和传递,起着承上启下的作用。当植物受到外界信号的刺激,比如射线、病原体、干旱等,静止状态的MAPK、MAPK激酶(MAPK kinase,MAPKK)和MAPKK激酶(MAPK kinase kinase,MAPKKK)会发生改变,产生顺序磷酸化反应,从而激活下游靶标基因的表达,对基因的表达、植物的生长和发育及对环境的适应进行调控[2]。在拟南芥中,可以通过4-甲氧基吲哚基-3-甲基硫代葡萄糖苷(4-methoxyindolyl-3-methylglucosinolates,4MI3G)的生物合成和积累,提供MAPK介导的信号通路来响应非生物胁迫[3]。
在拟南芥中MKK9-丝裂原活化蛋白激酶6(mitogen-activated protein kinase 6,MAPK6)级联系统通过调控Rubisco活化酶(rubisco activator,RCA)、质体核糖体蛋白1 (plastid ribosomal protein 1,PRPS1)、叶绿体分裂蛋白(chloroplast division protein,Ftsz2-2)和微管蛋白(tubulin,TOR2)的磷酸化在盐胁迫响应中发挥功能[1]。据报道,MKK9作为盐处理拟南芥愈伤组织中积累H2O2的正因子,与丝裂原活化蛋白激酶3/6(mitogen-activated protein kinase 3/6,MAPK3/6)形成通路在增强呼吸作用中起关键作用[4];
轻度渗透胁迫激活了MKK9-MAPK3/6级联信号系统,促进其在盐胁迫响应中发挥功能[5];
MKK9-MAPK3/6级联反应的激活促进了拟南芥幼苗的磷吸收[6];
核酮糖-1,5-二磷酸羧化酶/加氧酶合成因子22(ribulose-1,5-bisphosphate carboxylase/oxygenase assembly factor 22,RAF22)-MKK7/MKK9-MAPK3/MAPK6形成一个完整的过程级联,可能通过参与蔗糖的分解代谢调控拟南芥的生长[7];
磷脂酸(phosphatidic acid,PA)能够同时结合MAPK6和MKK9,增强MKK9对MAPK6的磷酸化活性[8];
MKK9的组成型和诱导型过表达导致拟南芥叶片过早衰老,MAPK6作为MKK9的直接靶标,敲除MKK9或MAPK6后叶片衰老被延迟[9];
拟南芥中MKK9是MAPK级联信号途径MAPK激酶(mitogen-activated protein kinase kinase,MKKs)中D组重要成员,可以响应低氮、低磷的胁迫调控花青苷的合成[10]。而苹果中MKK9在氮胁迫下对花青苷合成的调控机制鲜有报道。
青岛农业大学苹果育种课题组于2017年选育的R1R6型[成髓细胞瘤域蛋白质(myeloblastosis domain protein,MYB)10启动子区域有1个重复序列为R1,6个重复序列为R6]红肉苹果新品种‘黛红’(品种权号为CNA20162427.9)果实为绿皮红肉,是鲜食与加工兼用、具有广阔推广前景的红肉苹果新品种[8]。本试验以红肉苹果‘黛红’和‘新疆4号’为试材,提取成熟果实果皮果肉RNA,构建红肉苹果cDNA文库,利用实验室前期扩增得到的MdMKK9为诱饵筛选文库。通过酵母双杂交验证及体内双分子荧光互补(bimolecular fluorescence complementation,BiFC)方法进行MdMKK9互作蛋白验证,为红肉苹果花青苷生物合成调控机理提供一定的理论参考。
1.1 材料
1.1.1 试验材料
供试材料为10年生红肉苹果‘黛红’和‘新疆4号’,定植于青岛农业大学现代农业科技服务中心,砧木为八棱海棠。采集果实膨大期和成熟期的‘黛红’和‘新疆4号’果皮及果肉,置于-80 ℃保存备用。
1.1.2 试验药品
1.1.3 试验仪器
PCR仪(TaKaRa公司 TP-600)、植物活体荧光仪(法国Vilber公司 Newton 7.0)、电泳仪(上海艾研生物科技有限公司 PowerPac-3000)、凝胶成像系统(美国ProteinSimple公司 FluorChem R)、恒温培养箱(上海一恒科学仪器有限公司 DHP-9052)及摇床(广州市深华生物技术有限公司 IS-RDV1)等。
1.2 试验方法
1.2.1 红肉苹果cDNA文库构建
取适量果皮/果肉在液氮中迅速研磨成粉末,加入β-巯基乙醇及裂解液SL等试剂,对果皮果肉进行RNA提取,将检测合格的RNA样品送上海欧易生物医学科技有限公司进行cDNA文库构建,经单检混检均合格后得到红肉苹果cDNA酵母文库。
1.2.2 筛选基因载体构建
根据pGADT7载体图谱,按照同源重组方法设计MdAER、MdDFR、MdNRT2.7含XhoI酶切位点的引物,MdWRKY71、MdWRKY56、MdWRKY55、MdWRKY48、MdMYB87、MdANR、MdAlaAT2、MdMYB114、MdMAPK3含BamHI酶切位点的引物(见表1)。

表1 载体构建所用引物序列Table 1 The primers for construct vector
1.2.2.1 载体单酶切试验
文化不是虚的,强化企业文化建设,要塑造的是大家在一起做一番事业是意义和兴趣所在。企业文化更多的是组织对员工的承诺,同时也是员工对组织的承诺。其实是一种精神锁定,是一种心灵契约的锁定。员工在这种组织氛围下,无形之中会做出一种承诺,组织中的一群人都是这样的,如果自己将来不是这样,自己会受煎熬。同时,企业文化最重要的且最终是一种选择机制。企业文化不是解决所有的问题,不是所有人都会这样,而是尽量减少未来动荡的风险。
将上述反应体系,12 000 r/min涡旋离心1 min,充分混匀,37 ℃金属浴单酶切3~6 h,回收目的条带。

表2 载体单酶切反应体系Table 2 Carrier single enzyme digestion reaction system
1.2.2.2 同源重组连接试验
将上述反应体系手动轻轻吸打混匀,以12 000 r/min离心1 min后将反应液收集至管底。50 ℃金属浴15~30 min,降至4 ℃或立即置于冰上冷却。反应产物转化大肠杆菌DH5α感受态,挑取LB+Amp板上单克隆摇菌,菌液经交叉引物PCR筛选后进行测序,测序成功后命名为pGADT7-MdAER,pGADT7-MdDFR,pGADT7-MdNRT2.7,pGADT7-MdWRKY71,pGADT7-MdWRKY56,pGADT7-MdWRKY55,pGADT7-MdWRKY48,pGADT7-MdMYB87,pGADT7-MdANR,pGADT7-MdAlaAT2,pGADT7-MdMYB114,pGADT7-MdMAPK3将质粒与菌液于-80 ℃备用。

表3 同源重组连接反应体系Table 3 Homologous recombination linkage reaction system
1.2.3 酵母双杂交试验
1.2.3.1 Y2H Gold感受态的制备
用酵母膏蛋白胨葡萄糖腺嘌呤培养基(yeast extract peptone dextrose adenine medium,YPDA)活化酵母菌,30 ℃培养3 d。将单菌落点到含有3 mL YPDA培养基中。于250 r/min摇床上,在30 ℃下震荡培养8~12 h后,取适量菌液加入到50 mL新的YPDA培养基中。摇床振荡培养16 h至OD600=0.2。培养好的菌液用离心机7 000 r/min离心5 min,倒掉上清液,用100 mL YPDA进行重悬,吸打混匀,30 ℃孵育5 h至OD600=0.5。将摇好的菌液分装到50 mL灭菌离心管中,7 000 r/min离心5 min后弃上清液,加入30 mL无菌去离子水吸打重悬。将重悬液在7 000 r/min下离心5 min,弃上清液,用1.5 mL 1.1×TE/LiAC重悬液重悬。把上述细胞悬浮液转移到两个1.5 mL的离心管中,以12 000 r/min离心15 s后弃上清液。用600 μL 1.1×TE/LiAC重悬。
1.2.3.2 Y2H Gold感受态转化
将Bait质粒pGBKT7-MdMKK9质粒与Prey质粒pGADT7-MdCHS、pGADT7-MdSPX3按以下步骤共转Y2H Gold感受态细胞。
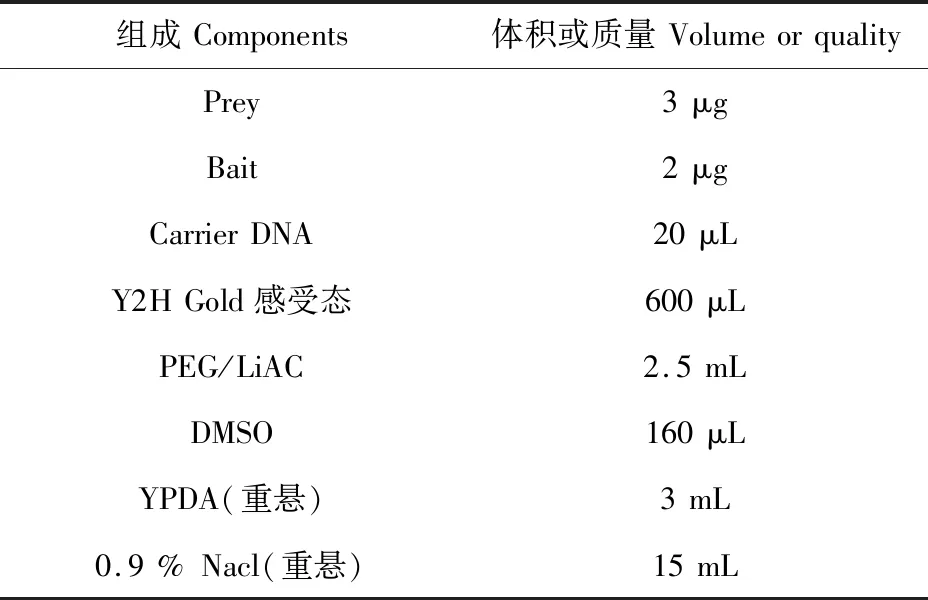
表4 酵母感受态转化体系Table 4 Yeast competent transformation system
1.2.3.3 质粒共转Y2H Gold感受态细胞
将上述重悬液均匀涂在SD/-Leu-Trp培养基上,30 ℃培养3~5 d。分别设置阳性(pGBKT7-53+pGADT7-T)与阴性(pGBKT7-Lam+pGADT7-T)对照。分别挑取对照与试验组培养基上的单菌落,用灭菌ddH2O稀释10倍,取8 μL点于SD-Ade-His-Leu-Trp/X-α-Gal/AbA培养基上,观察菌落颜色变化。

表5 酵母感受态转化体系Table 5 Yeast competent transformation system
1.2.4 BiFC验证互作
将无MdMKK9终止密码子的蛋白质编码区(coding sequences,CDS)克隆到pCAMBIA1300-nLUC载体中,并将MdSPX3克隆到pCAMBIA1300-cLUC载体上,将两种重组质粒转化大肠杆菌DH5α,送公司测序成功后,转化根癌农杆菌GV3101中。菌液于28 ℃摇床,摇至OD600=0.5~0.8,离心后用重悬液重悬,将nLUC-MdMKK9+cLUC-MdSPX3、nLUC-MdMKK9+cLUC、nLUC+cLUC-MdSPX3重悬液均匀混合,注射本生烟,培养2~3 d,使用活体荧光仪检测萤光素酶(luciferase,LUC)表达情况。
2.1 红肉苹果cDNA文库构建及筛库
MdMKK9在红肉苹果、尤其是本试验主要研究品种‘黛红’中发挥功能的互作蛋白还未可知。因此以‘黛红’和‘新疆4号’2个时期果皮果肉为试材提取RNA(图1A),由上海欧易公司经过RNA的单检及混检合格后构建红肉苹果cDNA文库,并进行互作蛋白的筛选验证。
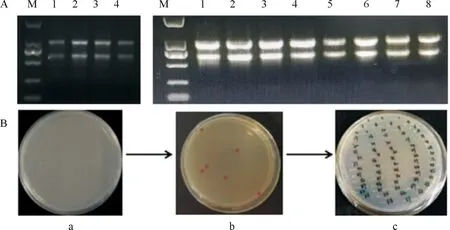
注:A.红肉苹果cDNA文库构建提取RNA;
M.DL2 000 Maker;
1-4(左图),‘黛红’果皮果肉RNA;
1-8(右图),‘新疆4号’果皮果肉RNA;
B.红肉苹果cDNA文库筛库过程;
a.将共转酵母菌液涂于SD-Trp-Leu+300 ng/μL AbA培养基;
b.SD-Trp-Leu+300 ng/μL AbA培养基长出单克隆菌斑;
c.SD-Ade-His-Trp-Leu+300 ng/μL AbA+X-α-gal显色反应。图1 红肉苹果cDNA文库构建及筛库Fig.1 Construction and screening of cDNA library of red flesh apple
如图1B,将pGBKT7-MdMKK9为诱饵蛋白,与红肉苹果cDNA文库质粒进行共转Y2H Gold感受态,涂于SD-Trp-Leu+300 ng/μL AbA培养基长出单克隆菌斑,并于SD-Ade-His-Trp-Leu+300 ng/μL AbA+X-α-gal培养基进行显色反应。
2.2 单克隆PCR及测序分析
2~3 d后,在SD-Ade-His-Trp-Leu+300 ng/μL AbA+X-α-gal培养基上得到400个蓝色菌斑,挑取菌斑,根据菌斑直径大小用灭菌双蒸水稀释10~20倍,取1~2 μL稀释液进行PCR筛选。选择单片段插入PCR产物20 μL进行测序分析。
将测序结果与NCBI数据库进行序列比对分析,最终选择了10个候选基因进行互作鉴定:花青苷合成相关基因ANR、DFR;
氮素相关基因AER、AlaAT2及NRT2.7;
抗逆相关基因WRKY71、WRKY48、MYB87、MYB114、MAPK3。以‘黛红’cDNA为模板克隆分别得到MdANR、MdMdAER、MdAlaAT2、MdDFR、MdNRT2.7、MdWRKY71、MdWRKY48、MdMYB87、MdMYB114、MdMAPK3。

注:M.DL2000Marker;
1-24.SD-Ade-His-Trp-Leu+300 ng/μL AbA+X-α-gal培养基上得到菌斑稀释后PCR产物电泳。图2 候选克隆PCR验证Fig.2 PCR validation of candidate clones
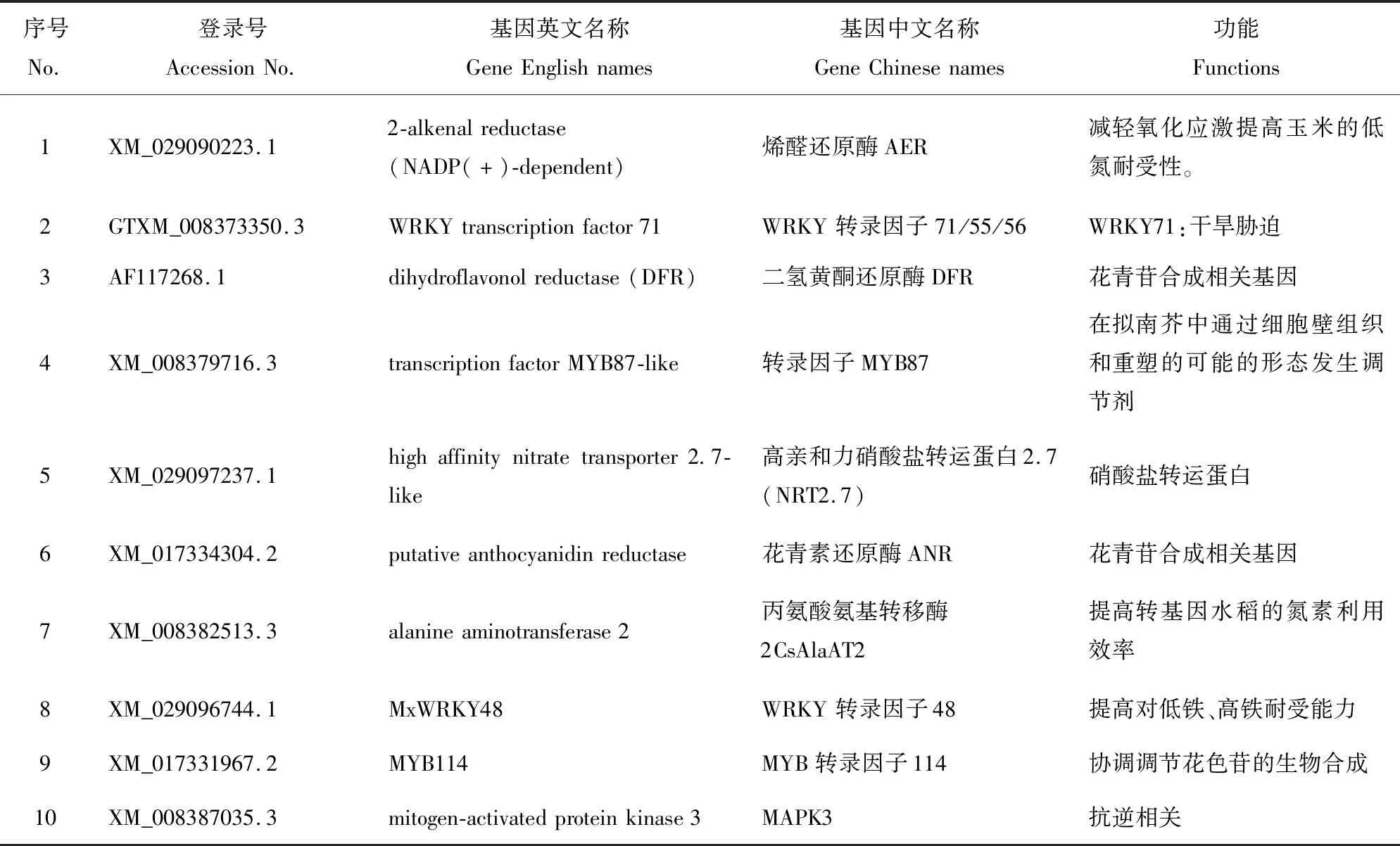
表6 互作蛋白功能注释Table 6 Functional annotation of the interacting protein
2.3 酵母双杂交验证互作
构建AD-MdANR、AD-MdMdAER、AD-MdAlaAT2、AD-MdDFR、AD-MdNRT2.7、AD-MdWRKY71、AD-MdWRKY48、AD-MdMYB87、AD-MdMYB114、AD-MdMAPK3载体,分别与MdMKK9共转于Y2H Gold感受态,涂于SD-Trp-Leu+300 ng/μL AbA与SD-Ade-His-Trp-Leu+300 ng/mL AbA+X-α-gal培养基上。选取AD-MdANR+BD、AD-MdMdAER+BD、AD-MdAlaAT2+BD、AD-MdDFR+BD、AD-MdNRT2.7+BD、AD-MdWRKY71+BD、AD-MdWRKY48+BD、AD-MdMYB87+BD、AD-MdMYB114+BD、AD-MdMAPK3+BD、BD-MdMKK9+AD作为对照。
由图3结果显示,在SD-Trp-Leu+300 ng/μL AbA培养基上,AD-MdANR、AD-MdMdAER、AD-MdAlaAT2、AD-MdDFR、AD-MdNRT2.7、AD-MdWRKY71、AD-MdWRKY48、AD-MdMYB87、AD-MdMYB114、AD-MdMAPK3与MdMKK9、BD共转菌斑均生长。但在SD-Ade-His-Trp-Leu+300 ng/μL AbA培养基上只有AD-MdMYB87+BD-MdMKK9菌斑生长且颜色变蓝,其他菌斑均未生长及变色。试验结果表明,只有MdMYB87可以与MdMKK9互作。

图3 酵母点对点验证Fig.3 Point-to-point validation of yeast
2.4 BiFC验证
为进一步验证MdMYB87与MdMKK9的互作情况,构建了pCAMBIA1300-nLUC-MdMKK9、pCAMBIA1300-cLUC-MdMYB87载体,进行BiFC试验,通过观察LUC表达情况得知MdMKK9与MdMYB87在体内互作。
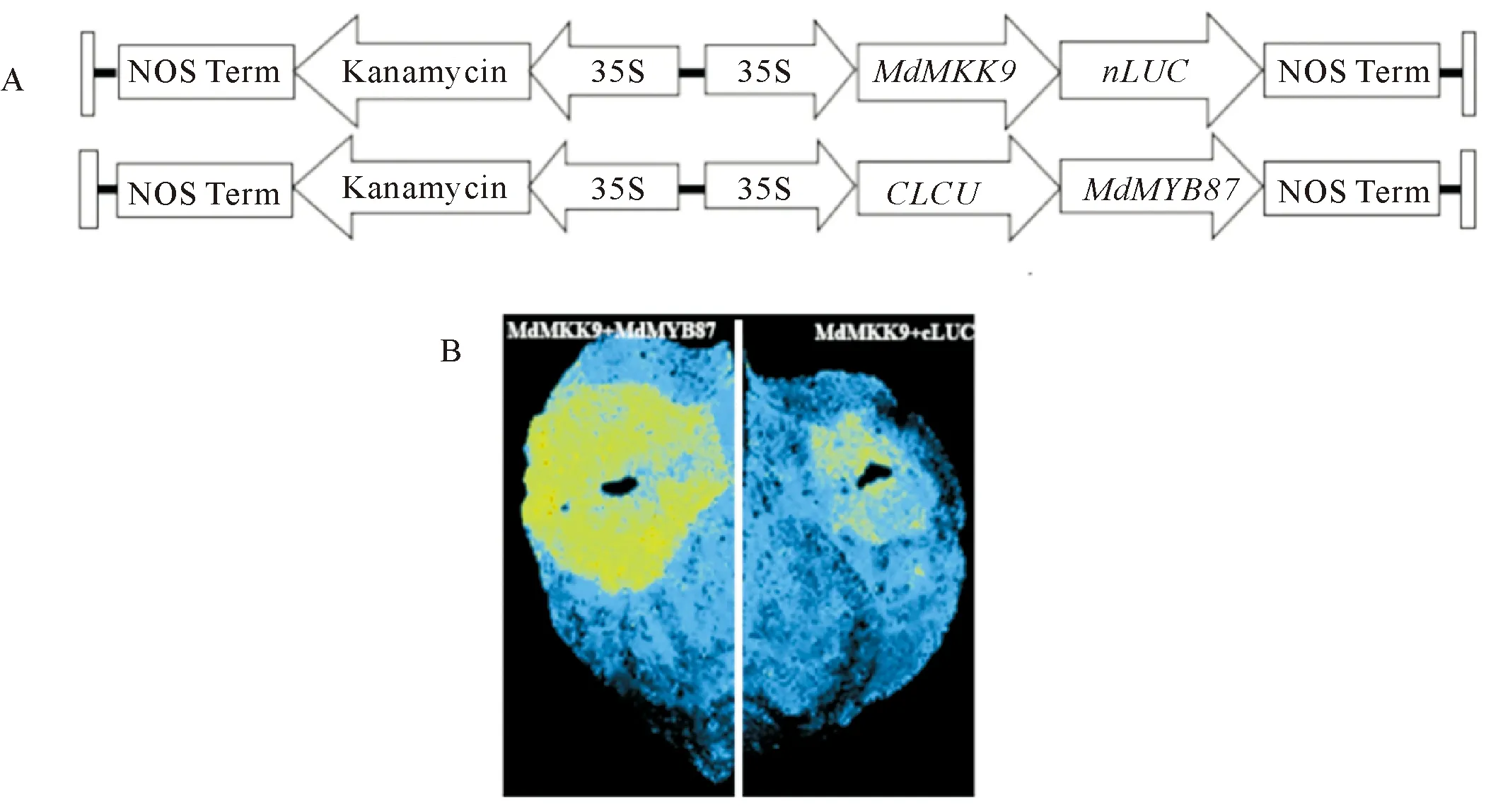
注:A.pCAMBIA1300-nLUC-MdMKK9、pCAMBIA1300-cLUC-MdMYB87载体构建图;
B.LUC基因表达情况。图4 BIFC验证MdMKK9与MdMYB87互作Fig.4 The interaction between MdMKK9 and MdMYB87 by BIFC verification
蛋白质的互作及互作研究具有十分重要的生物学意义,在细胞接受内外源信号、通过信号途径调节基因表达、保持植物生物学特性的过程中,蛋白质都占有重要地位,且大部分的蛋白质是通过与伴侣分子或其他的蛋白质复合物一起发挥作用。因此,我们需要通过酵母双杂技术筛选出蛋白质的互作蛋白,用来更好的研究蛋白质单体、复合物的功能以及细胞生物学活性。
本试验以MdMKK9作为诱饵蛋白,提取了‘黛红’及‘新疆4号’果实RNA构建红肉苹果cDNA文库,通过筛选MdMKK9在红肉苹果中的互作蛋白,发现了与抗逆相关的基因MdMYB87。在近期的研究中:张顺仓等[11]研究发现SmMYB87蛋白在细胞核和细胞膜上均有分布,在根、茎、叶和花中均有表达,该基因通过介导丹参对生物或非生物胁迫的响应来调控其有效成分的积累;
嵌合抑制因子MYB87的组成性表达会抑制多个器官的纵向、径向生长,通过细胞壁组织和重塑调控拟南芥的形态发生[12];
生物序列的相似性搜索(basic local alignment search tool,BLAST)分析表明,在拟南芥中,包含MYB36、MYB37、MYB38、MYB68、MYB84和MYB87的R2R3Myb基因亚群与番茄(blind-like1,Bl)基因的相似性最高,番茄Bl基因编码一种R2R3Myb转录因子,该转录因子在营养和生殖发育期间对腋生分生组织起始的调节中起着核心作用[13];
对拟南芥根细胞进行转录组分析,得出了形成层表达的转录因子编码基因包括短营养期(short vegetative period,SVP)、花瓣损失(petal loss,PTL)和MYB结构域87(MYB87),在拟南芥中,位于形成层的维管干细胞在次生生长过程中不断自我更新和分化,干细胞持续进行细胞增殖,并产生传导性组织,持续的干细胞活动依赖于细胞增殖和细胞分化之间的严格平衡[14]。
通过酵母双杂交及BiFC最终验证MYB87与MdMKK9互作。但MdMYB87对MdMKK9在响应逆境胁迫花青苷合成的途径中的调控功能尚不清楚,有待进一步展开研究。
猜你喜欢 菌斑文库红肉 用生命保卫中央文库的陈为人湘潮(上半月)(2022年7期)2022-12-06伊朗红肉月产量增长11%今日农业(2022年13期)2022-11-10大学生牙龈炎龈上菌斑的微生物群落昆明医科大学学报(2022年2期)2022-03-29加工红肉才是影响健康的“罪魁祸首”中老年保健(2021年6期)2021-11-30新西兰红肉出口超10亿新元,中国为最大出口目的地今日农业(2021年13期)2021-11-26新西兰红肉出口保持强劲态势今日农业(2021年16期)2021-11-26专家文库猪业科学(2021年3期)2021-05-21点亮经典文学之灯——“百年文库”第一辑隆重推出少先队活动(2020年12期)2021-01-14关于推荐《当代诗坛百家文库》入选诗家的启事中华诗词(2019年1期)2019-11-14龈下菌斑对慢性牙周炎患者种植体周围边缘骨吸收的影响中外医学研究(2017年6期)2017-03-30推荐访问:蛋白 激酶 活化