玉米C型胞质不育系S37-2败育的生物学与生理生化机制分析
来源:优秀文章 发布时间:2023-04-25 点击:
郑 冉,吕 丹,武清贵,邸晓红,朱通通,邱冠杰,罗红兵,2,*
(1.湖南农业大学 农学院,湖南 长沙 410128;
2.湖南省玉米工程技术研究中心,湖南 长沙 410128)
不育化制种是当前玉米制种的一种重要方式[1]。细胞质雄性不育(CMS)是由细胞核和细胞质基因共同控制的雄性不育,可实现不育系、保持系和恢复系的三系配套。玉米CMS分为T型、C型和S型3类,其中C型不育系败育较彻底,育性表现稳定,在玉米不育化制种中具有巨大的潜力[2]。
玉米CMS的发生过程存在异常的生理代谢,对其生理生化特性进行研究可将玉米遗传特性与个体表型联系起来,可为揭示玉米雄性不育的发生机理提供参考[3]。营养物质的亏缺被认为是CMS产生的一个重要原因[4]。研究发现,玉米雄性不育材料中的可溶性蛋白、可溶性糖和脯氨酸等物质的含量少、代谢水平低[5-8]。株系的正常发育都会伴随着营养物质的积累与合成,因此,不育材料中营养物质含量的降低,必然会造成代谢紊乱,从而导致不育。活性氧(ROS)的积累也被认为是CMS产生的原因之一,当ROS过多时,会促使丙二醛(MDA)积累,从而引起相关组织的生理生化紊乱[9-11]。段俊等[12]研究表明,玉米不育系叶片和雄穗中的ROS含量高于保持系;
邓杰[5]对玉米细胞质雄性不育系的研究发现,不育系对OH·的清除速率和MDA含量高于保持系;
张勤[7]试验发现,玉米不育系叶片中MDA、H2O2和O2-含量均高于同期的可育系。过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)是活性氧清除系统和抗氧化系统中重要的酶。研究结果显示,不育材料中SOD和CAT的活性低于可育材料[13-16],而POD活性高于或低于可育材料,差异较大[17]。由以上研究可以推论,CMS与植物体内较高的活性氧水平,以及由此产生的较高程度的膜脂过氧化有密切关系。本研究以玉米雄性不育系S37-2为研究材料,通过对其表型性状、理化指标的分析,初步探究其雄性不育的生理生化机理,以期为该不育系的实践应用奠定基础。
1.1 试验材料
供试品种为C型胞质雄性不育系S37-2及其保持系B37-2,由湖南农业大学农学院罗红兵教授提供。
1.2 试验设计
试验材料分别于2021年3月和8月种植于湖南农业大学耘园试验基地。试验土壤为红色土,肥力水平中等。试验采用小区种植,不育系S37-2与保持系B37-2均设置3次重复,每个试验小区种植2行,行长3 m,每行10穴,穴距30 cm,行距50 cm。
玉米追肥于第7叶完全展开时进行,施用尿素300 kg·hm-2。分别在苗期(一叶一心期,幼苗)、拔节期(由上至下第5片叶)、抽雄期(雄穗尖端露出顶叶3~5 cm时,最上部展开叶)、散粉期(最上部展开叶)、成熟期(最上部展开叶)取相应部位的叶片。将叶片冷冻于液氮中带回,保存于-80 ℃冰箱。
1.3 指标测定
1.3.1 主要农艺性状调查
于成熟期,每小区随机选取10株植株定株,调查其株高、穗位高、茎粗、茎节数、叶片数、穗位叶叶长、穗位叶叶宽、雄穗分枝数、雄穗长等性状。
1.3.2 花器官形态特征调查
于散粉期,对各个材料花粉的育性进行调查。随机剥取散粉的雄穗中部花药,选取测量10个花药的长度和横截面直径,对比S37-2和B37-2的区别。
1.3.3 不同发育期叶片生理指标测定
分别选取不同发育时期的叶片,测定其可溶性糖、可溶性蛋白、游离脯氨酸、可溶性淀粉、丙二醛含量,以及CAT、SOD、POD活性。具体地,采用硫酸蒽酮法测定可溶性糖含量[18],采用考马斯亮蓝比色法测定可溶性蛋白含量[19],采用茚三酮比色法测定游离脯氨酸含量[20],采用碘显色法测定可溶性淀粉含量[21],采用紫外吸收法测定CAT活性[13],采用NBT光还原法测定SOD活性[22],采用愈创木酚法测定POD活性[23],采用硫代巴比妥酸(TBA)法测定MDA含量[24]。每个样品重复测定3次。
1.4 数据分析
采用Excel 2020软件整理数据,采用SAS 9.4软件进行方差分析,对有显著(P<0.05)差异的,采用Duncan法进行多重比较。
2.1 主要农艺性状比较
对比不育系S37-2与保持系B37-2的主要农艺性状差异(表1)。方差分析结果表明,S37-2的穗位叶叶长、叶宽均显著高于B37-2,除此以外的其他性状差异均不显著。
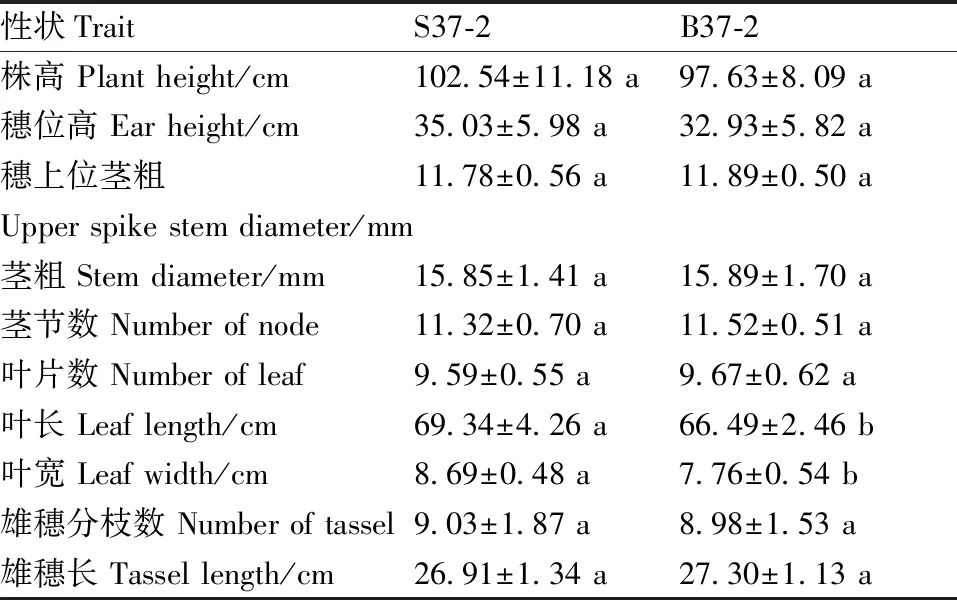
表1 S37-2与B37-2的农艺性状比较
2.2 花药表型特征比较
对比S37-2与B37-2的花药发现,S37-2的花药干瘪皱缩,B37-2的花药形状饱满(图1)。对花药表型性状进行方差分析,结果表明,B37-2花药的长度和横截面直径均显著高于S37-2(表2),S37-2花药的长度仅为B37-2的68.52%,横截面直径仅为B37-2的40.00%。

图1 不育系S37-2(左)与保持系B37-2(右)的花药形态Fig.1 Anthers morphology of sterile line S37-2 (left) and maintainer line B37-2 (right)

表2 B37-2与S37-2的花药性状比较
2.3 不同发育期叶片营养物质含量比较
随着玉米植株的发育,可溶性蛋白含量在S37-2和B37-2叶片中均呈先降后升再降的趋势(图2)。S37-2的可溶性蛋白含量在散粉期达到最大值,B37-2的可溶性蛋白含量在出苗期达到最大值,两者的可溶性蛋白含量均在成熟期最低。S37-2叶片中的可溶性蛋白含量在出苗期和拔节期显著低于B37-2,在抽雄期和散粉期显著高于B37-2,成熟期两者差异不显著。

同一时期柱上无相同字母的表示差异显著(P<0.05)。下同。Bars marked without the same letters at the same growth stage indicate significant difference at P<0.05. The same as below.图2 S37-2和B37-2叶片中的营养物质含量比较Fig.2 Comparison of nutrients content in leaves of S37-2 and B37-2
从出苗期到成熟期,S37-2和B37-2叶片的游离脯氨酸含量变化趋势基本一致,但S37-2在散粉期达到最大值,为158.57 μg·g-1。除拔节期和成熟期外,其他发育时期S37-2叶片中的游离脯氨酸含量均显著高于B37-2,分别是B37-2的1.24、1.13和1.57倍。
从出苗期到成熟期,S37-2和B37-2的叶片可溶性糖含量变化趋势一致,变化幅度较小,均随植株发育先降后升再降,均在散粉期达到最大值,分别为3.56%、3.15%。拔节期—成熟期,S37-2的可溶性糖含量均显著高于B37-2,分别为B37-2的1.13、1.03、1.17和1.05倍。
S37-2的叶片可溶性淀粉含量在各发育期变化幅度较小。B37-2的叶片可溶性淀粉含量随植株发育呈先升后降的变化趋势,在散粉期达最大值,并于成熟期下降。从出苗期到成熟期,S37-2的叶片可溶性淀粉含量均显著低于同时期的B37-2,分别是B37-2的63.84%、48.88%、46.12%、48.80%和52.16%。
2.4 不同发育期叶片酶活性比较
随着玉米植株的发育,S37-2与B37-2叶片的CAT活性均呈先降后升再降的趋势(图3)。S37-2叶片的CAT活性在散粉期最高,在拔节期最低;
B37-2叶片的CAT活性也在散粉期最高,但在抽雄期最低。S37-2叶片的CAT活性在各时期均显著低于B37-2,分别是B37-2的80.72%、80.47%、92.28%、81.55%和87.56%。

CAT、SOD、POD活性均以鲜重计。Activities of CAT, SOD and POD were measured based on fresh weight.图3 S37-2和B37-2叶片中酶活性与丙二醛(MDA)含量的比较Fig.3 Comparison of enzymes activities and malondialdehyde (MDA) content in leaves of S37-2 and B37-2
从出苗期到成熟期,B37-2叶片的SOD活性均呈上升的趋势,与B37-2相比,S37-2叶片的SOD活性在各时期都显著更低。
在玉米叶片的发育过程中,S37-2和B37-2叶片中的POD活性总体呈先升后降的趋势,最高值均出现在散粉期,分别为553.65、497.82 U·g-1(以鲜重计),最低值均出现在出苗期。S37-2叶片中的POD活性在各个时期均显著高于B37-2。
S37-2和B37-2叶片的MDA含量随玉米发育均呈现先降后升的趋势,在成熟期达到最大值。整个发育过程中,S37-2叶片的MDA含量与同时期的B37-2均无显著差异。
通过对S37-2和B37-2农艺性状的比较可知,S37-2的穗位叶叶长、叶宽显著大于B37-2,这可能是由于S37-2的生殖生长受阻,更多的营养物质用于叶片生长。S37-2的花药形状瘦小萎缩,饱满度差,花药的长度、横截面直径均显著低于B37-2,这与赵宇等[13]的研究结果一致。
植物在发育过程中需要大量的蛋白、淀粉、糖与氨基酸,当这些物质的合成和运输路径受阻时,植物雄穗花药的生长发育将会受到影响,甚至产生不育的现象[25]。营养物质的缺失会导致小麦[26]、大豆[27]等作物的小孢子败育。李玉青等[28]的研究表明,不育系中的可溶性糖、可溶性蛋白和脯氨酸含量较低。本研究发现,在S37-2与B37-2之间,其叶片的营养物质含量差异变化并没有特别明显的规律性,与其他雄性不育材料整个发育期营养物质均明显缺乏的现象不一致。推测这可能与该类型材料雄性不育的发生时期相关,也可能与试验材料的取样部位、试验环境等有关。S37-2叶片中的可溶性蛋白、游离脯氨酸和可溶性糖含量均在抽雄期与散粉期显著高于B37-2。据此推测,S37-2叶片中这3种物质的运输路径受阻,无法向雄穗提供足够的营养物质,从而影响小孢子的发育和育性表现,导致败育。这也进一步说明,玉米的雄性不育会受到物质代谢的影响。
玉米正常的生长发育需要植物抗氧化酶系统的调节,防止膜脂过氧化,维持动态平衡。前人研究发现,CMS花药败育过程中,CAT、SOD、POD等酶的活性存在不同程度的波动。柴伟国等[29]发现,辣椒不育系的花药POD活性明显高于保持系;
蒋会兵等[30]发现,厚轴茶不育株花蕾中的CAT、SOD活性显著低于可育株,POD活性高于可育株;
邓杰[5]、邹佳等[14]研究玉米雄性不育系发现,不育系中CAT、SOD活性低。本试验结果表明,在生长发育的各个时期,S37-2叶片中的CAT和SOD活性均显著低于保持系(B37-2),这与其他学者在小麦和水稻等雄性不育系上的研究[31-32]相似。同时,本研究发现S37-2的POD活性显著高于B37-2。这可能是不育株的自我保护反应,但也可能会造成花粉内营养物质的匮乏,最终导致雄性不育的发生[33-34]。
综上推测,S37-2叶片中的可溶性蛋白、游离脯氨酸和可溶性糖的运输路径受阻,以及CAT和SOD活性降低,可能是导致其雄性不育的重要原因。上述结果仅反映了玉米雄性不育系S37-2叶片发育过程中的生理生化变化趋势,其他方面还需进一步深入研究。
猜你喜欢散粉保持系花药高粱制种田存在的问题及对策研究园艺与种苗(2021年10期)2021-11-28略施“魔法” 妆容不融化北京青年周刊(2019年28期)2019-08-02大豆不育系及其同型保持系生殖生长期生理特性比较研究中国种业(2018年8期)2018-08-28天边的霞光,脸上的红晕优雅(2017年10期)2017-10-16生态因子对滇重楼花药开裂的影响广西植物(2016年10期)2016-11-11误把散粉当痱子粉环境与生活(2016年9期)2016-10-08杂交籼稻花药基部开裂性状的遗传研究西南农业学报(2016年4期)2016-05-17杂交稻三系配套的原理及功能研究种子科技(2015年8期)2015-05-24甘蓝雄性不育系PM·QM及其保持系的开花结实特性研究安徽农业科学(2015年13期)2015-01-13植物花药开裂机制研究进展中国蔬菜(2013年8期)2013-01-28推荐访问:生物学 生化 玉米推荐文章
- 2018年江西赣州市医疗急救中心招聘编外人员公告:赣州市医疗急救中心
- 升学宴策划活动方案|2018升学宴活动方案
- 小学六一游园活动总结 [小学六一游园活动策划]
- 2018年中国工商银行广东分行暑期实习生招聘岗位、报名时间:2018中国工商银行广西分行春招
- 澳洲留学八大名校排名申请条件_澳洲留学奖学金申请条件及时间
- [调工商档案介绍信范文] 工商档案查询介绍信
- 加拿大亲属移民政策最新更新|加拿大亲属移民条件
- 初一下册语文练习册答案人教版2018 2018人教版语文练习册答案
- 贵州贵阳房价2018 2018年贵州贵阳中医学院第二附属医院招聘方案
- 【2018广东省湛江市赤坎区审计局招聘公告】2018湛江市赤坎区教师招聘