不同类型调理剂对镉污染土壤修复效果和微生物群落的影响
来源:优秀文章 发布时间:2022-11-15 点击:
李奇,王艳红,李义纯,李林峰,唐明灯,艾绍英
(广东省农业科学院农业资源与环境研究所/农业农村部南方植物营养与肥料重点实验室/广东省养分资源循环利用与耕地保育重点实验室,广东广州510640)
【研究意义】土壤镉(Cd)污染严重威胁我国稻田安全生产。近年来,我国土壤Cd污染面积和强度呈不断增长的趋势,华南地区稻田土壤Cd污染更为普遍和严重(Yang et al.,2018;
Peng et al.,2019)。土壤原位钝化修复技术主要是通过添加化学调理剂改变Cd在土壤中的赋存形态、降低其生物有效性,从而使农产品达到安全生产的目的(刘晓月等,2018;
周利军等,2019)。目前,常用于Cd污染农田土壤修复的调理剂为各种硅钙物质(如硅酸钠、硅酸钙、石灰等)、有机废弃物(如畜禽粪便、有机堆肥)和微生物菌剂(如芽孢杆菌、地杆菌),这些不同类型的调理剂在改变土壤有机质和活性矿物含量、改善微生物活性、降低土壤Cd的生物有效性及减少作物可食用部分的累积量方面发挥着不同作用(Liu et al.,2018;
Hussain et al.,2020;
Albuquerque et al.,2021),但其在影响土壤微生物群落多样性方面的作用尚不清楚。近年来,调理剂作为一种有效的Cd钝化剂受到越来越多的关注,也非常适合于大面积中低度Cd污染农田土壤的修复,能更好地满足当前我国Cd污染土壤的治理与农产品安全生产的需求。因此,研究调理剂对Cd超标农田土壤修复效果和微生物群落的影响,对我国稻米安全生产和农田生态可持续发展具有重要意义。【前人研究进展】土壤微生物是维持土壤生物学特性的重要组成部分(Pan et al.,2020;
朱永官等,2021),其中微生物群落和多样性的变化可用来研究调理剂对重金属污染土壤的生态调节作用,判断调理剂对农田土壤生态安全的影响。微生物的群落结构和功能影响着土壤结构、氧化还原状态及酶活性等(Kasemodel et al.,2019;
褚海燕等,2020),从而改变Cd在土壤中的溶解度和生物有效性(Beattie et al.,2018;
Zhao et al.,2021)。Ali等(2011)研究发现,伯克霍尔德菌属可提高土壤Cd的生物有效性,并增加玉米和番茄植株对Cd的吸收。Suksabye等(2016)于Cd污染土壤中添加2%(v/v)的铜绿假单胞菌、枯草芽孢杆菌和球孢白僵菌,发现水稻籽粒中Cd含量分别降低74.16%、60.14%和44.84%,与其他菌株相比,经铜绿假单胞菌改良的土壤中稻谷Cd积累量最低,表明铜绿假单胞菌较其他菌株吸附更多的Cd。有研究表明,盆栽Cd污染土壤中添加50 mL CJW-1菌株(约10CFU/mL)40 d后,其对Cd的固定率可达56.10%(Peng et al.,2020)。随着新一代测序技术的发展,一些研究表明,Cd污染土壤中微生物群落随环境条件发生变化,但变形杆菌、厚壁菌、酸杆菌、放线杆菌和拟杆菌等被认为是Cd污染环境中持久存在的优势菌门,在调控Cd有效性方面发挥着重要作用(Zhang et al.,2016;
李海龙等,2018;
Fagnano et al.,2020;
Hao et al.,2021)。有研究报道,石灰配施5%的生物质炭,农田土壤中有效态Cd含量降低98.1%,土壤微生物基础呼吸强度和土壤微生物量碳含量显著增加(曾秀君等,2020)。施用海泡石、石灰和秸秆生物炭使得土壤微生物群落的多样性指数(ACE、Chao1和Shannon)增加,群落结构发生显著改变,变形菌门、厚壁菌门和拟杆菌门相对丰度有所增加,芽单胞菌门、绿弯菌门和螺旋体菌门相对丰度降低(兰玉书等,2020)。可见,土壤微生物群落结构的变化一定程度可有效反映添加调理剂对农田土壤生态的影响。【本研究切入点】目前,有关钝化剂对盆栽或小区试验土壤微生物群落结构的影响研究较多,但基于不同土壤活性组分的调理剂对田间污染土壤中微生物群落结构及其生态安全的影响鲜见研究报道。【拟解决的关键问题】通过田间试验,研究添加不同活性组分调理剂(矿物型、有机型和微生物型)对土壤基本理化性质、Cd生物有效性、微生物群落结构和农田生态安全的影响,探究不同类型调理剂作用下土壤环境因子与微生物群落结构的关系,评估其对农田土壤生态环境质量的影响,以期为应用不同土壤活性组分调理剂原位修复和治理Cd污染农田土壤提供科学依据。
1.1 试验区概况
于2020年3—7月,在广东省兴宁市寺岗村水稻田(东经115°69′,北纬24°34′)开展田间试验。试验前,土壤的理化性质为:pH 5.21,有机碳31.49 g/kg,碱解氮181.6 mg/kg,有效磷7.26 mg/kg,速效钾49.65 mg/kg,阳离子交换量13.67 cmol/kg。土壤中总Cd含量为0.70 mg/kg,超过《土壤环境质量农用地土壤污染风险管控标准(试行)》(GB 15618—2018)中Cd的风险筛选值(水田,5.5<pH≤6.5,0.4 mg/kg),但低于污染风险管制值(水田,5.5<pH≤6.5,2.0 mg/kg),说明在该区域进行水稻种植,存在糙米Cd含量超过《食品安全国家标准 食品中污染物限量》(GB 2762—2017)中Cd限量值(0.2 mg/kg)现象。
1.2 试验材料
(1)矿物型调理剂:本团队自主研发产品,主要成分为硅酸钙、熟石灰、硫酸钾、无水硫酸镁和九水合硝酸铁。本团队以往的研究发现,这种改良剂会通过提高土壤pH、促进无定形铁矿物及铁硫化物对Cd吸附或共沉淀,从而影响土壤中有效态Cd含量(李义纯等,2019,2020)。(2)有机型调理剂:主要成分为蚕沙和烟梗(有机质含量≥47%),N+PO+KO≥35%,购自广东植物龙生物技术股份有限公司。(3)微生物型调理剂:金葵子复合微生物肥,其有效活性菌数≥0.2亿个/g(枯草芽孢杆菌、地衣芽孢杆菌和米曲霉),N+PO+KO≥8%,购自佛山金葵子植物营养有限公司。3种调理剂总Cd含量均低于《肥料中砷、镉、铅、铬、汞生态指标》(GB/T 23349—2009)中Cd的限量值。
1.3 试验设计
在上述区域进行施用不同土壤活性组分调理剂的田间试验,设3个处理:T1,矿物型调理剂;
T2,有机型调理剂;
T3,微生物型调理剂。每处理重复3次,面积为1/3 ha,3种调理剂田间施用量均为3 t/ha。同时设空白对照(CK),除不添加调理剂外,其他田间管理措施均保持一致。为防止各处理间相互影响,在原有田埂的基础上进行加固。为使材料与土壤混合均匀,3种土壤调理剂在水稻种植前7 d结合整地翻耕一次性撒施。
1.4 样品采集
试验土壤在水稻成熟期采集,采集时间为2020年7月。每处理采用五点取样法采集土壤0~20 cm样品和水稻样品。土样除去根茎等杂质,装入100 mL离心管,置于4℃车载冰箱中,在24 h内带回实验室-80℃保存;
部分土壤风干过筛后进行土壤理化性质分析。
1.5 分析方法
1.5.1 土壤理化性质参照鲍士旦(2000)的方法对土壤理化性质进行测定,pH采用电位法测定;
有机碳(SOC)采用重铬酸钾—油浴法测定;
全氮(TN)采用凯氏定氮法测定;
全磷(TP)采用NaOH熔融—钼锑抗比色法测定;
全钾(TK)采用NaOH熔融—火焰光度法测定;
碱解氮(AN)采用碱解扩散法测定;
有效磷(AP)采用碳酸氢钠提取—钼锑抗比色法测定;
速效钾(AK)采用乙酸铵提取—原子吸收法测定;
阳离子交换量(CEC)采用乙酸铵交换法测定。
1.5.2 土壤有效态Cd和糙米Cd测定采用DTPA浸提法提取和测定有效态Cd含量(GB/T 23739—2009)。将稻谷样品晾晒至恒重,再对其进行脱粒、脱壳、研磨和消解(Chen et al.,2018),然后采用石墨炉原子吸收光谱仪(PE AA600)测定消解液中Cd含量,同时利用植物标准样品GBW10048(GSB-26)(中国地质科学院地球物理地球化学勘查研究所IGGE生产)对整个消解和分析测定过程进行质量控制,标样中Cd的回收率为95.6%。
1.5.3 微生物群落分析利用DNA提取试剂盒(FastDNASpin Kit for Soil,MP Biomedicals,美国)进行基因组DNA抽提,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA。将DNA样品送至上海美吉生物医药科技有限公司进行测序分析。对测序得到的原始数据(Raw data)进行质控检测、过滤,得到有效数据(Clean data)。首先使用Qiime(1.7.0,http://qiime.org/)按如下标准去除低质量序列:(1)平均质量分数<20分以及长度<50 bp的序列;
(2)Barcode错配1个碱基及以上的序列;
(3)引物中错配>2个碱基的序列。然后使用FLASH(1.2.7,http://ccb.jhu.edu/software/FLASH/),基于重叠序列长度>10 bp,重叠区域不允许有模糊碱基的标准,对序列进行拼接。最后用mothur去除嵌合体。
所得到的高质量序列按照97%的相似性阈值划分分类操作单元(Operational taxonomic unit,OTU),并去除只含有1条序列的OTU(singleton)。为保证不同样品测序深度一致,对所有样品的序列按相同的序列数进行抽平,抽平后的数据用于后续注释及统计分析。选取每个OTU中最长的序列作为该OTU的代表序列,使用BLASTn在NCBI非冗余数据库(nr)中对代表序列进行检索和比对,以1×e作为值的最小阈值,去除>1×e的参考序列,然后从剩下的序列中选取得分(Total score)最高的参考序列对该OTU进行注释。若所有检索到的参考序列所对应的值均大于1×e,则表示该代表序列尚无同源序列,分类位置无法确定。
2.1 不同调理剂对土壤理化性质及有效态Cd和糙米Cd含量的影响
从表1可看出,添加调理剂可显著提高土壤pH(增幅0.11~0.24)(<0.05,下同),有效缓解土壤酸化的情况。矿物型(T1)、有机型(T2)和微生物型(T3)调理剂处理后土壤有机碳含量显著增加,增幅分别为32%、47%和21%。矿物型和有机型调理剂显著增加土壤CEC值,微生物型调理剂则显著降低CEC值。3种调理剂显著提高土壤全氮含量,其中有机型调理剂的效果最明显,增幅为37%。矿物型调理剂显著增加土壤全磷含量,增幅为27%,但有机型调理剂显著降低其含量。对土壤全钾而言,微生物型调理剂显著提高其水平,增幅为2%,而矿物型和有机型调理剂显著降低其含量。可见,3种调理剂在改善土壤全量养分上的作用不一。对土壤速效养分而言,3种调理剂均显著提高土壤碱解氮、有效磷和速效钾含量,改善土壤营养状况,其中有机型调理剂对土壤碱解氮和有效磷的改善最明显,分别增加26%和188%,微生物型调理剂在提高土壤速效钾含量上的作用最佳,增幅达63%。
不同调理剂处理后土壤DTPA提取态Cd和糙米Cd含量如图1所示。与CK相比,矿物型、有机型和微生物型调理剂均显著降低土壤DTPA提取态Cd含量,降幅分别为13%、48%和28%(图1-A)。对糙米Cd含量而言,矿物型、有机型和微生物型调理剂均显著降低其含量,其中矿物型和微生物型调理剂的效果相当,最大降幅达42%,有机型调理剂对糙米Cd含量的降低幅度最小(26%)(图1-B)。可见,调理剂施用后土壤有效态Cd和糙米Cd含量均有明显下降,其中有机型调理剂对土壤有效态Cd降低最显著,但对糙米Cd含量的降低效果不佳。
2.2 施加不同调理剂后土壤中细菌和真菌的群落组成
通过高通量测序,不同样品的序列数在25089~53584条,对数据进行抽平处理,使每个样品序列数均为25089条,然后按照97%的序列相似性进行OTU聚类并产生OTU表。使用BLASTn在NCBI非冗余数据库(nr)对每个OTU的代表序列进行检索和比对,选取得分(Total score)最高的参考序列对该OTU进行注释。
在细菌门水平上,施用不同调理剂的土壤中仅有4.1%~5.5%的序列未检索到相似序列或相对丰度低于1%,剩下的序列由放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、氯曲菌门(Chloroflexi)、酸杆菌门(Acidobacteriota)、厚壁菌门(Firmicutes)、硝基螺菌门(Nitrospirota)、脱硫菌门(Desulfobacterota)、粘球菌门(Myxococcota)、拟杆菌门(Bacteroidota)、芽单胞菌门(Gemmatimonadota)、扁平菌门(Planctomycetota)、MBNT15、Sva0485和疣微菌门(Verrucomicrobiota)组成(图2-A)。其中放线菌门和变形菌门的相对丰度在所有调理剂处理中均较高。方差分析表明,3种调理剂处理均显著降低土壤中脱硫菌门和拟杆菌门的相对丰度;
其中矿物型和有机型调理剂提高放线菌门和芽单胞菌门的相对丰度,微生物型调理剂提高酸杆菌门的相对丰度。

该地区土壤样品的真菌一共注释到6个门,占检索序列的87%~90%,由子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、被子菌门(Mortierellomycota)、罗兹菌门(Rozellomycota)、壶菌门(Chytridiomycota)和球囊菌门(Glomeromycota)组成。其中子囊菌门的相对丰度在所有处理中均占绝对优势(42%~57%)(图2-B)。方差分析结果表明,矿物型调理剂显著增加担子菌门的相对丰度,降低子囊菌门的相对丰度;
而有机型调理剂显著增加被子菌门相对丰度,降低罗兹菌门的相对丰度;
微生物型调理剂显著增加子囊菌门的相对丰度,降低罗兹菌门的相对丰度。
2.3 细菌和真菌群落结构组成对不同调理剂处理的响应
通过对所获得的OTU进行热图聚类分析发现,不同调理剂处理之间其OTU的相对丰度明显不同(图3)。然而,细菌的聚类分析与真菌有相似规律,即均有分化成矿物型调理剂和其他调理剂2个类别,再进行有机型和微生物型调理剂处理的分类。这也暗示了施加矿物型调理剂对细菌和真菌物种组成的影响与其他2种调理剂处理方式显著不同。
不同调理剂处理下细菌和真菌群落结构的非度量多维尺度分析(Non-metric multidimensional scaling,NMDS)排序结果(图4)显示,调理剂类型对这两类微生物的群落结构变化均有显著影响,样品均能按照调理剂类型聚类在一起,而在不同调理剂处理之间分开。相似性分析(Analysis of similarities,ANOSIM)是一种非参数检验分析方法,常被用来检验2组或多组间差异是否显著大于组内差异,从而判断分组类型是否有意义。结果显示,细菌ANOSIM的=0.9290(<0.01),而真菌ANOSIM的=0.7716(<0.01)。此外,处理间的分组椭圆也明显分开,均可认为CK和3种调理剂处理之间的细菌和真菌群落结构存在明显差异。


2.4 环境因子对细菌和真菌群落结构组成的影响
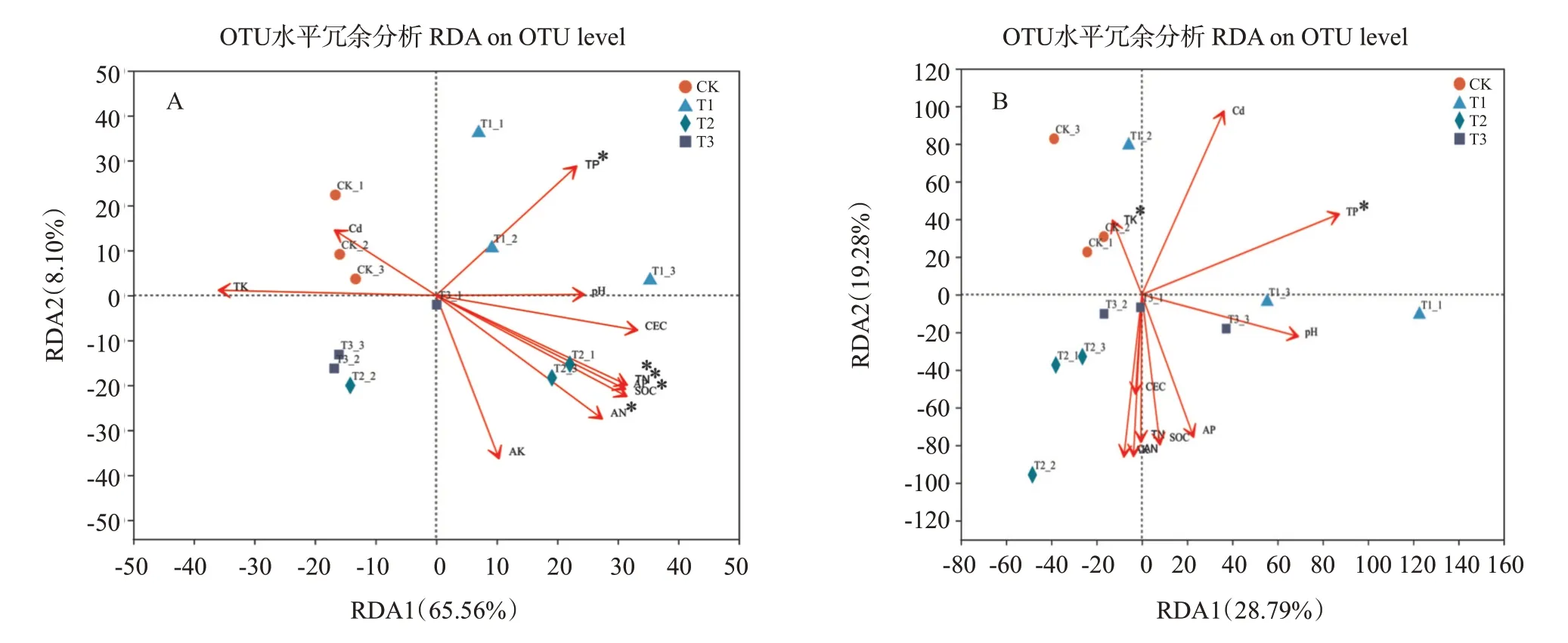
在生态学领域中,冗余分析(Redundancy analysis,RDA)是常用的约束排序分析方法,其目的是分析“解释变量(一般为环境因子矩阵)”对“响应变量(一般为物种矩阵)”的影响情况。通过RDA分析发现所测得的理化因子对细菌和真菌微生物群落的解释度分别为73.66%和48.07%,其中轴1分别能解释65.56%和28.79%(图5)。对细菌的群落结构而言,施用矿物型调理剂能在轴1上与CK分开,在轴2上与另外2种调理剂处理分开;
此外,有机型和微生物型调理剂均能在轴2上与CK分开(图5-A)。对真菌的群落结构而言,有机型和微生物型调理剂均能在轴2上与CK分开(图5-B)。999次蒙特卡洛检验显示,全磷是显著影响细菌和真菌微生物群落结构共同的主要环境因子。此外,土壤有机碳、全氮、碱解氮、有效磷和全钾分别是显著影响细菌和真菌微生物群落结构的其他环境因子。土壤全磷与施用矿物型调理剂在细菌群落呈现出明显的正比关系,而与微生物型调理剂处理呈反相关;
土壤中的有机碳、全氮、碱解氮、有效磷含量则明显与施加有机型调理剂呈正相关。
2.5 土壤Cd有效性与微生物群落的关系
土壤DTPA提取态Cd、糙米Cd含量与微生物群落的相关分析结果(图6)表明,DTPA提取态Cd含量与细菌或真菌群落的相关性均不显著(>0.05,下同)。糙米Cd含量与细菌群落的变形菌门、脱硫菌门和拟杆菌门呈显著正相关,其相关系数分别为0.61、0.83和0.60,其与真菌群落的相关性均不显著。



3.1 土壤有效营养成分的提高
不同活性组分调理剂通过改变土壤理化性质和生物特性来有效改善肥力和土壤质量,从而提高作物产量(Kavitha et al.,2018),土壤中碱解氮、有效磷和速效钾常被选作评价土壤肥力的参数。与对照相比,土壤碱解氮、有效磷和速效钾分别增加了12%~26%、118%~188%和28%~63%。在已有报道中,矿物基土壤调理剂含硅、钙、镁、钾等养分,可促进水稻生长,对水稻增产有重要作用(Chen et al.,2020a)。有机型调理剂属富碳物质,有机碳含量高,在土壤中施入可直接提高土壤碳储量。本研究使用的有机型调理剂主要为蚕沙肥,其有机质含量超47%,对土壤速效养分的提升最明显。李北齐等(2011)对生物有机肥进行室内和大田试验均表明,生物有机肥能明显增加土壤pH,同时提高有机质、速效磷和速效钾含量,本研究结果与其结果一致。这可能是由于有机质不仅能吸附和固定土壤中的NO、NH和PO,减轻气态氮损失,还可减少养分浸出,提高土壤肥力(Yang et al.,2018)。有研究表明,微生物肥料施入土壤后,能迅速生长繁殖并形成有益微生物菌群,通过植物—微生物共生体系,溶解释放土壤养分(Bargaz et al.,2018;
鲁洪娟等,2019)。谢修鸿和李玉(2010)通过盆栽试验研究发现,菌糠菌肥增加土壤速效氮、磷、钾含量,尤以速效磷、钾增幅较大,分别增加9倍和4倍。然而,本研究中微生物型调理剂处理对土壤碱解氮和速效磷的提升效果最差。推测这种不同可能是由于在田间条件下添加的外源微生物存活率较低导致。此外,施入微生物菌肥能增强作物抵抗逆境的能力,从而达到增产提质的效果(葛均青等,2003;
Ai et al.,2015;
Kour et al.,2020)。这是因为其含有的有益微生物菌群、活性物质、营养物质及多种微量元素,能改善土壤微环境、激活酶活性、促进作物对营养物质的吸收利用,进而提高作物产量。
3.2 稻田土壤Cd和糙米Cd含量的关系
本研究结果发现,土壤有效态Cd与糙米Cd含量的相关性不显著,值得注意的是,有机型调理剂处理下,两者呈相反关系。以往的研究认为,稻田土壤和稻米Cd含量的关系复杂,因外界环境不同,存在正相关性和非正相关性(田茂苑等,2019;
黄雁飞等,2020)。彭华等(2013)、张建辉等(2015)的研究显示,土壤有效态Cd与稻米Cd含量的相关系数均相对较低。然而,有研究发现,稻米富集Cd受土壤Cd形态变化的显著影响,稻米Cd含量与土壤可交换态Cd呈极显著正相关(潘杨,2015;
王梦梦等,2018)。造成以上相关性差异的可能原因有:(1)水稻自然生长环境的复杂性及土壤其他理化性质的差异性较大,研究中未考虑到土壤类型、气候、灌溉及施肥农艺等因素的影响(龚伟群等,2006;
范美蓉等,2012)。(2)不同于盆栽试验的高相关性系数和极显著相关性,大田环境下由于土壤类型和理化性质具有一定的空间异质性,作物生长环境条件不均一(赵科理,2010;
廖启林等,2015),导致与盆栽试验不同的研究结果。总体而言,稻米Cd与土壤Cd含量关系复杂,除呈正相关性外,还存在其他关系。因此,降低稻米Cd含量的关键是了解稻田土壤与稻米Cd含量之间的相关关系,但就目前公布的研究数据而言,多为局部区域数据,如Fang等(2014)通过92份稻米样品说明我国主要种植水稻区域Cd的健康风险,王梦梦等(2018)用60个点位研究14个乡镇稻田和稻米Cd含量,用于说明二者的定量关系,这些数据难以全面、准确地揭示稻米与土壤Cd的关系。进一步研究应有针对性、全面性地对稻田土壤和稻米Cd含量关系开展协同研究,同时通过开展关于多因素对稻米Cd含量的影响,建立多因素与土壤有效态Cd和稻米Cd含量之间的线性关系,为Cd污染稻田修复治理提供理论依据。
3.3 土壤微生物群落的改善
作为土壤的活跃组分,微生物区系组成及微生物数量与土壤的理化性质变化密切相关。现有研究显示,通过改变土壤的理化性质从而影响根系微生物的群落结构,例如改变根际土壤结构、保水性、营养物质等,形成有利于某些菌种生长或不利于某些菌种生长的环境条件,从而改变土壤微生物的群落结构(Chen et al.,2020b;
Hermans et al.,2020)。本研究利用高通量测序,分析3种土壤调理剂处理下农田土壤的细菌和真菌群落结构及多样性,以及其与土壤理化特性的相关性。结果显示,调理剂处理下细菌和真菌群落丰富度及多样性明显高于对照。本研究土壤中细菌群落主要隶属于放线菌门、变形菌门、氯曲菌门、厚壁菌门和酸杆菌门,其中,放线菌门和变形菌门在所有样品中均占有绝对优势;
对真菌群落而言,主要由子囊菌门、担子菌门、被子菌门、罗兹菌门、壶菌门和球囊菌门构成,其中子囊菌门占主要地位。这与大多数农田土壤环境中微生物多样性研究结果(兰玉书等,2020;
曾鹏等,2021;
Pei et al.,2021)一致。
方差分析结果表明,3个调理剂处理均显著降低土壤中脱硫菌门和拟杆菌门的相对丰度;
矿物型和有机型调理剂显著提高土壤中放线菌门和芽单胞菌门的相对丰度,微生物型调理剂显著提高土壤中酸杆菌门的相对丰度。糙米Cd含量与微生物群落的相关分析结果显示,糙米Cd含量与细菌群落的变形菌门、脱硫菌门和拟杆菌门呈显著正相关。这些结果意味着糙米Cd含量的明显下降与调理剂处理均显著降低脱硫菌门和拟杆菌门的相对丰度有关。此外,兰玉书等(2020)研究表明,在轻度Cd污染水稻田中施加钝化剂后土壤中的变形菌门最高增加24%,然而不少研究报道变形菌门对重金属污染不敏感,能在重金属浓度较高的环境中生存(Xu et al.,2016;
Wang et al.,2019)。一般认为,酸杆菌门嗜酸,可在寒冷和酸性等不良环境中生存,参与到植物残体的降解和铁循环中,有利于改善土壤的生态微环境(Pankratov et al.,2011;
Mao et al.,2012)。本研究结果显示,添加微生物型调理剂后土壤pH升高,酸杆菌门相对丰度有所增加。也有研究发现在中性、碱性的环境中检测出酸杆菌基因序列,并表示酸杆菌门的相对丰度与土壤中多种环境因子相关(Navarrete et al.,2013),推测可能与酸杆菌门对土壤生态功能响应存在差异有关(王光华等,2016)。这似乎意味着微生物型调理剂在调控和改善土壤菌群结构方面有着较强的应用潜力。在以往的研究中,施用螯合铁肥,复合碳酸钙、钙镁磷肥和含硫有机物,以及复合炉渣和生物炭处理后,土壤细菌的多样性指数提高2.55%~3.21%(周雅心等,2020;
Chen et al.,2020c)。可见,添加土壤调理剂可维持甚至提高污染土壤中正常微生物群落的多样性和丰富度,对农田土壤的生态安全无明显影响。然而,基于RDA和NMDS分析表明调理剂处理可显著影响土壤微生物群落的β-多样性,且不同活性组分调理剂的影响差异显著。因此,添加不同组分调理剂均可改变土壤微生物群落的组成和结构。
土壤调理剂处理下农田土壤中细菌群落的分布与环境因子有很大关系,本研究发现全磷是影响细菌和真菌群落结构的重要环境因子,此外土壤有机碳、全氮、碱解氮、有效磷和全钾是影响微生物群落组成的其他环境因子。有研究表明,土壤中pH、有机碳、总氮、有效磷和电导率对细菌群落组成影响较大(丁新景等,2018)。屈忠义等(2021)研究发现,土壤中添加不同改良剂后,细菌和放线菌数量均与土壤pH和电导率呈负相关,与土壤有机质、水解氮、速效磷和速效钾含量呈正相关,并指出可将土壤细菌和放线菌数量作为评价土壤健康状况的重要生物指标。施用高碳基和高钙镁调理剂后,土壤速效钾是烟田土壤细菌群落变异的主导性因子(谢仕祺等,2021)。也有研究表明,pH是影响微生物群落的关键因素,兰玉书等(2020)报道,pH与微生物群落功能多样性指数显著相关,说明该因素对微生物群落影响较大,而土壤营养物质对微生物群落的影响并不明显。但在本研究中并未得到此结论,推测该现象是由于土壤中pH相对较低,因此可在农业种植过程中适当提高土壤pH,更有利于土壤微生物活性和健康的群落结构组成。可见,不同调理剂/钝化剂处理后土壤的细菌群落结构及多样性差异的理化因子不尽相同。
在粤东地区Cd污染稻田添加土壤调理剂可提高土壤pH,增加土壤有机质,改善土壤的有效养分状况,显著降低土壤有效态Cd和糙米Cd含量,其中矿物型和微生物型调理剂对糙米Cd含量的控制效果最佳。总体上施用调理剂可提高土壤肥力和土壤微域环境的健康状况。磷素是影响细菌和真菌群落结构共同的重要环境因子。从降低土壤有效态Cd和糙米Cd含量及对土壤微生物群落结构影响方面综合考虑,微生物型土壤调理剂修复效果最佳,降低水稻对Cd吸收的同时改善土壤菌群结构。
猜你喜欢 糙米群落调理 轻度人为干扰下贵州道地药用植物艾纳香群落特征研究山地农业生物学报(2022年3期)2022-05-13缺氮胁迫对小麦根际土壤微生物群落结构特征的影响*土壤学报(2022年1期)2022-03-08人教版生物必修3第4章第4节群落的演替教学设计学校教育研究(2020年7期)2020-04-09种群和群落及生态系统考题分析中学生理科应试(2017年6期)2017-09-27改善糙米食用品质的研究进展食品界(2017年3期)2017-04-19给宝宝引入糙米的黄金期亲子(2014年4期)2014-05-10糙米减肥餐饮食科学(2009年5期)2009-05-14自制“糙米茶”,排毒又减肥祝你幸福·午后版(2008年10期)2008-09-20流产者的饮食调理祝您健康(1992年6期)1992-12-28身体瘦弱者的饮食调理祝您健康(1989年2期)1989-12-30推荐访问:群落 微生物 调理