江鳕生化遗传学特性及群体遗传结构初步分析
来源:优秀文章 发布时间:2022-12-02 点击:
张林,甘金华,何力,张涛,周剑光
(中国水产科学研究院长江水产研究所,农业农村部淡水鱼类种质监督检验测试中心,湖北 武汉 430072)
近年来,水产养殖在很大程度上是依靠消耗大量生产要素资源、自然资源来维持的,造成渔业资源衰退。面对种质资源遭受到的威胁,我们对中国大部分鱼类的种质情况尚不明了。种质鉴定能反映鱼类种群资源状况,查明种群的遗传结构和变异情况,确定原种保护和开发利用措施[1]。
江鳕(LotaLota),又名鲇鱼、山鲶鱼、山怀子,属于鳕形目,鳕鱼科,江鳕属(Lota),鳕科鱼类中唯一的淡水鱼种。该鱼种属北极淡水复合体鱼种,是典型的冷水性鱼类,具有酷寒期于冰下生殖洄游,并在水温接近 0 ℃时产卵的特殊习性。该鱼种广泛分布在北美和欧亚的一些内陆水域和海湾,属于高纬度水域珍贵经济鱼类,因具有肥大的肝脏和美味的雄性精巢而著称[2]。中国仅新疆的额尔齐斯河、东北的黑龙江水系及鸭绿江上游水域有该鱼种的分布。该鱼种可作为防冻、抗寒及低温状态下生命活动等方面研究的特定物种。
目前关于该鱼种相关的研究很少,仅见黑龙江水系及鸭绿江上游的江鳕生物学方面的报道,包括江鳕对环境的适应性、影响江鳕徊游的因素、年龄、生长、食性、繁殖、形态学及亚种的分类[3-6],江鳕作为野生鱼种常被用来研究环境污染状况的指标。对额尔齐斯河水域江鳕的研究非常少,更未对江鳕进行系统的种质研究和种质标准的制定。近年随着江鳕捕获量的增加,野外水域中该鱼种越来越少,因此对该鱼种的种质资源开展生化遗传学、分子生物方面的系统研究,对该鱼种的开发利用以及资源保护具有非常重要的意义。本研究利用聚丙烯酰胺凝胶电泳技术对乳酸脱氢酶、乙醇脱氢酶、苹果酸脱氢酶、异柠檬酸脱氢酶4种同工酶在江鳕的肌肉、肝脏、心脏、脾脏、肾脏和眼睛等6 种组织中的分布、活性及遗传变异进行了比较分析,旨在了解4种同工酶在江鳕6种不同组织中的差异性表达,解析江鳕额尔齐斯河群体的遗传学结构,为该鱼种野生种质资源保护及江鳕属鱼类的分类、物种鉴定和系统进化研究提供理论依据。
1.1 试验动物
试验动物江鳕30尾(雌雄各15 尾),由浙江海洋大学于2017年 7月采自新疆额尔齐斯河水系的布尔津河流域,样品体长在 20.3~22.5 cm 之间,体重为 230~246 g,江鳕经充氧活体送至实验室养殖,暂养于室内2.0×2.0×1.8 m循环水鱼池中一周后开始实验。
1.2 样品采集及样本制备
样品采集:将鱼池中的10尾鱼(雌雄各5尾)搬运至实验室,剪鳃放血处死,将鱼体表面水分擦干后解剖,分别取背部肌肉、肝脏、心脏、脾脏、肾脏和眼睛等6种组织,编号后放入相应的塑料袋,于-80 ℃低温冰箱保存备用。
样本制备:取0.3~0.5 g冷冻组织样品,使用生理盐水将血液清洗干净,剔除表面的杂质,按1∶3的比例(m/v)加入新鲜灭菌蒸馏水,于干净的匀浆器中充分匀浆后以12 000 r/min低温离心30 min分离上清,将上清液转入新离心管中,重复 2~3次,直至上清液澄清为止,整个操作过程需在低温下进行。
1.3 同工酶的凝胶电泳分析
同工酶采用聚丙烯酰胺(PAGE)梯度凝胶电泳分析。聚丙烯酰胺梯度凝胶电泳方法参照Nei等[8]的报道,并加以改进。聚丙烯酰胺梯度凝胶电泳所采用4.0%的浓缩胶和7.0%的分离胶,电泳时采用pH 8.3的Tris-甘氨酸电泳缓冲液,加样前,先用50 mA电流进行30 min恒流预电泳以除去胶中杂质,然后向每个加样孔中加入20 μL制备好的样品,用280 mV电压进行约10 min电泳,待样品出凝胶孔后,将电压调至220 mV进行电泳,待上样溴酚蓝指示剂电泳至平板底处时,结束电泳,整个电泳过程均在4 ℃下进行。
1.4 凝胶的染色和脱色
电泳结束后,将胶片从玻璃板中剥离,放入37 ℃恒温箱中预热的染色液进行染色,染色至所有条带清晰时终止。各种同工酶的染色过程参照Shaklee和Nei等[7-8]的报道。染色好的胶片放入2.5%冰乙酸中进行脱色,脱色后清晰显示的胶片,放入保存液中进行固定。
1.5 图谱分析与数据处理
固定好的凝胶照片用凝胶成像系统(Bio-RAD Gel DOCTMXR+)进行拍照并分析,并用专用水溶性玻璃纸将胶片固定在玻璃板上,置于阴凉处晾干,以利于长期保存。电泳图谱中各种同工酶的缩写、命名、编号、酶谱分析、基因位点、等位基因和基因型的命名参照Nei和Hofmann等的报道[8-9]。基因位点的命名以向阳极迁移的速度快慢,采用数字依次编号。依据电泳结果判断每种同工酶的基因型,根据基因型计算每个等位基因的基因频率。采用多态位点比例和平均杂合度2个指标来衡量遗传变异的程度。平均杂合度、多态位点比例、遗传距离等参数的计算参照Nei[8]和日本水产资源保护协会报道的方法[9-10],即实测等位基因的出现频率小于或等于0.99时,可视为多态。参照杨元吴[11]的方法对数据进行统计处理。
2.1 江鳕同工酶的组织特异性表达
2.1.1 乳酸脱氢酶的组织差异性表达
乳酸脱氢酶为四聚体酶,在江鳕中由 5个基因位点编码,5个基因位点均为多态。在眼睛中出现5条酶带,多态性明显。LDH1在脾脏、肾脏中不表达,在其他组织中微弱表达,LDH1条带在肌肉中的颜色最深,活性最强;
LDH2在肝脏中不表达,在其他5个组织中均有分布;
LDH3在各组织中均有表达,条带清晰,在肝脏中表达较弱,其余组织中表达均较强,LDH2、LDH3在心脏中的颜色最深,活性最强;
LDH4、LDH5仅在肝脏及眼中有表达。江鳕LDH 同工酶的电泳结果见图1。

图1 江鳕LDH同工酶的电泳图谱1-3:肌肉,4-6:肝脏,7-9:心脏,10-12:脾脏,13-15:肾脏,16-18:眼睛。下同。Fig.1 Electrophoretograms for LDH isozyme of Lota lota1-3:muscle, 4-6:liver, 7-9:heart, 10-12:spleen, 13-15:kidney,16-18:eye.The same below.
2.1.2 乙醇脱氢酶的组织差异性表达
乙醇脱氢酶是一个分子量为80 kDa的二聚体,主要催化乙醇转化为乙醛。江鳕的 ADH由5个基因位点编码,ADH1、ADH2、ADH3为多态,ADH4和ADH5为单态。ADH1仅在肝脏和肾脏中表达,ADH2在肌肉、肝脏和肾脏中表达,ADH3在心脏中表达最丰富,在肌肉和肝脏中无表达,在其他组织中有微弱表达,ADH4除在肝脏中表达较弱,在其他组织中活性均较强,其中心脏与眼中活性最高,ADH5仅在肝脏、眼中有微弱的表达。江鳕ADH 同工酶的电泳结果见图2。

图2 江鳕 ADH同工酶的电泳图谱Fig.2 Electrophoretograms for ADH isozyme of Lota lota
2.1.3 苹果酸脱氢酶的组织差异性表达
苹果酸脱氢酶为二聚体酶,包括上清液型(s-MDH)和线粒体型(m-MDH)两种形态,二者不能相互转化形成异二聚体。江鳕的MDH由两个基因位点编码,均为多态,MDH-1在除脾脏外其他组织均可检测到,MDH2在所检测的组织中均可检测到。江鳕MDH同工酶的电泳结果见图3。

图3 江鳕MDH同工酶的电泳图谱Fig.3 Electrophoretograms for MDH isozyme of Lota lota
2.2 江鳕群体遗传结构参数
通过分析江鳕的肌肉、肝脏、心脏、脾脏、肾脏和眼睛等6种不同组织中的LDH、ADH和MDH等3种同工酶代表性条带,初步了解了江鳕群体遗传学参数(表1)。经统计共记录基因位点12个,10个基因位点有多态,多态位点的基因频率为0.125~0.875,多态位点比例为83.33%;
等位基因数为22,位点平均有效等位基因数为1.833 3;
平均观测杂合度为0.423 4,平均期望杂合度为0.310 9,Hardy-Weinberg 遗传偏离指数 D为0.361 9。
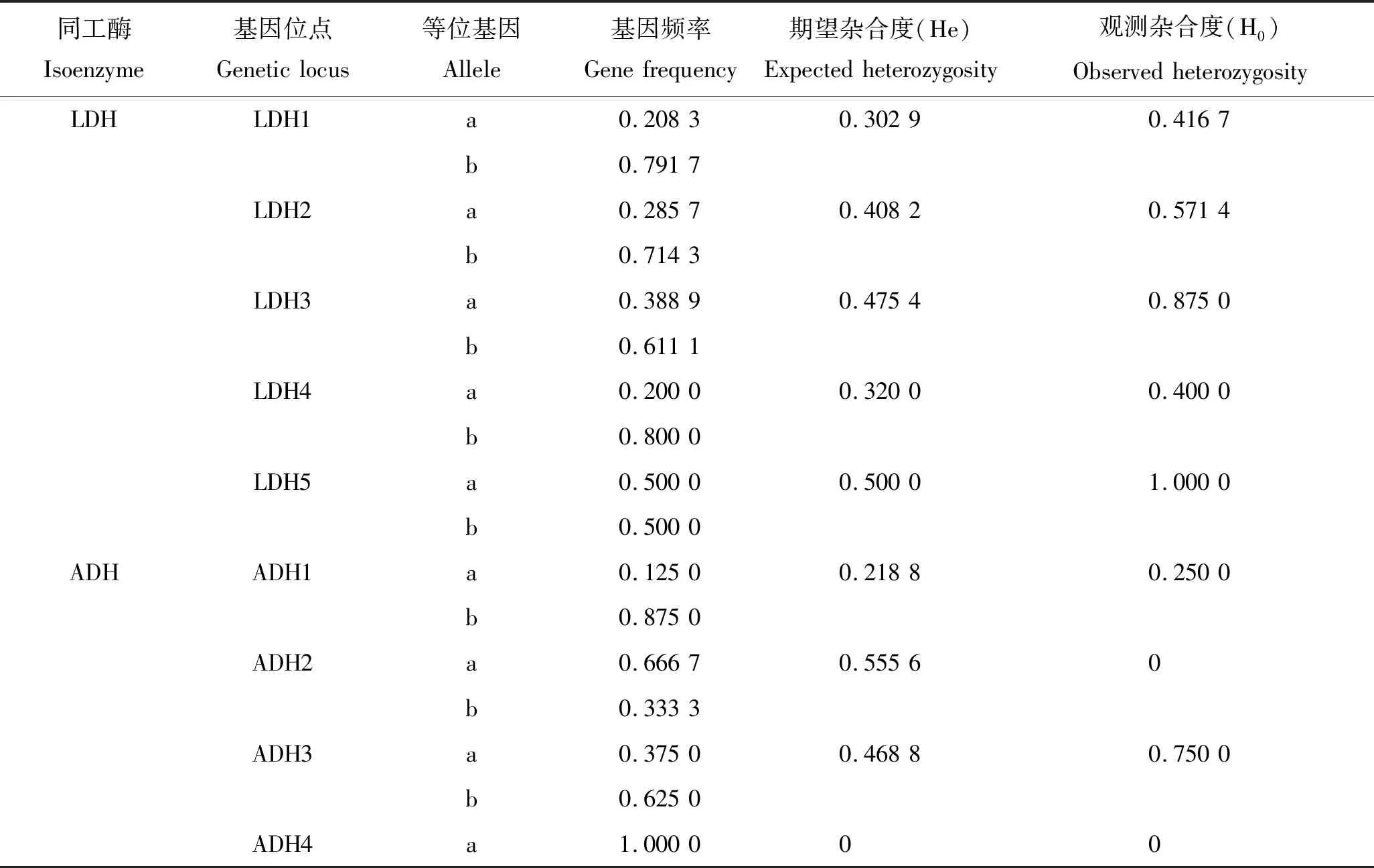
表1 江鳕群体的遗传学结构Tab.1 Genetic structure of Lota lota
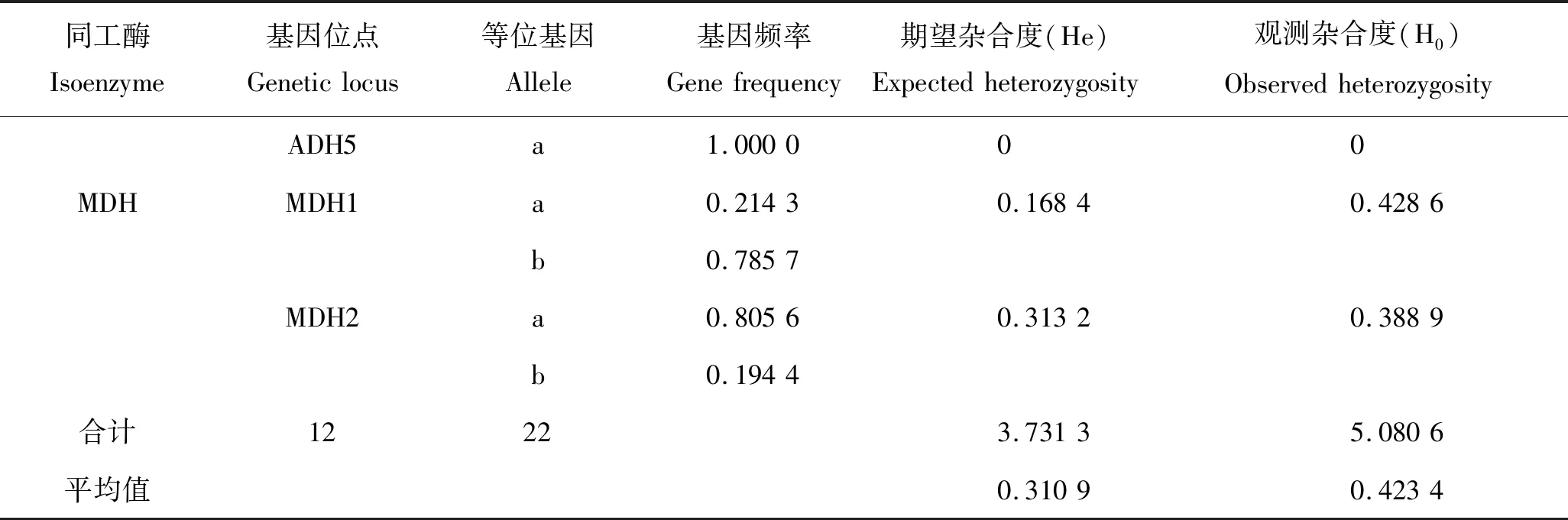
续表1,Tab.1 Continued
2.3 江鳕生化遗传学标记
首先随机取5尾鱼,进行6种组织常见同工酶(LDH、ADH、MDH、EST、IDHP)筛查,然后取10尾健康江鳕对筛选的单态同工酶进行验证,以初步确定反映江鳕种质的生化遗传特性。结果表明:江鳕肌肉组织异柠檬酸脱氢酶(IDHP)是单态,且稳定表达,可作为江鳕生化遗传学标记。江鳕肌肉组织IDHP电泳结果见图4。
图4 江鳕肌肉组织IDHP电泳图谱1-10泳道分别表示不同个体的酶谱。Fig.4 Electrophoretogram of IDHP isozyme expressed in muscle of Lota lota1-10 showed the zymograms of different individuals.
3.1 江鳕同工酶表达的组织特异性分析
通过对江鳕的肌肉、肝脏、心脏、脾脏、肾脏和眼睛6种组织的LDH、ADH、MDH、EST、IDHP 5种同工酶进行电泳分析,发现同工酶表达具有显著的组织特异性。因江鳕许多组织及个体酯酶多次未能检出,因此本研究仅选择LDH、ADH和MDH 3种经典的同工酶酶谱初步分析江鳕的遗传学结构。研究发现LDH、ADH、MDH 3种同工酶在心脏中均有高水平进行表达,其次是眼睛、肝脏、肌肉,肾脏和脾脏中各同工酶的表达逐渐减弱,多态水平高。同时对多种组织进行IDHP电泳,发现只有肌肉组织是单态,表达1条酶带,可以作为江鳕特有的种质生化遗传标记。而其他组织IDHP表达不清晰,拖带严重。
LDH一般由A和B两种不同多肽链组成的同源或异源四聚体。本研究中LDH1由A基因编码形成四聚体,基本上在无氧条件下起作用,这说明江鳕肌肉无氧代谢较为旺盛,与其作为冷水底栖凶猛肉食鱼类的习性相适应[12-13]。LDH3由B基因编码形成四聚体,在有氧组织中表现出很高的活性,这与心脏催化乳酸氧化形成丙酮酸进入三羧酸循环相一致。在心脏中的LDH3、ADH3、ADH4 和 MDH2特异性高表达,进一步说明了心脏无可替代的代谢功能。江鳕LDH的组织分布证明了江鳕很可能存在编码 LDH-C的基因。已有研究表明,LDH-C的四聚体仅在哺乳动物的精子以及高等鱼类的肝和眼中检测到[14-16]。本研究中发现江鳕眼睛的LDH同工酶有5条特异性多态条带,说明由LDH-C基因编码,与已报道的研究结论相符,该结果也表明江鳕在进化上处于相对高级的阶段。分析原因可能是江鳕在淡水环境中处于食物链顶端,所面临的选择压力较大,因此其进化速度相比其他高等鱼类更为快捷。
3.2 江鳕群体的遗传结构分析
种群的遗传结构能够反映物种的进化过程,多态位点比例(P)和平均杂合度(H)指标是衡量鱼类群体生化遗传变异和种质资源状况的重要指标[15-18]。多态位点比例会因鱼种或同种鱼的不同种群而出现差异,本研究中得到的 P 值及 He均较高,表明江鳕额尔齐斯群体在生化遗传变异上也处于较高水平,当然也可能由于本研究所选代表性酶种类较少,其次IDHP、EST各组织中检测到杂合子的情况较少或未检出。因此,如果多种酶进行综合考虑,其P值和He值可能较本研究中常见的有代表性的3种酶谱所得出的数据小些,但即使这样,额尔齐斯河江鳕群体的遗传变异性高于鱼类的平均水平。
遗传偏离指数(D) 也是全面反映群体的遗传平衡状况的重要指标。D值接近零则基因的分布就接近平衡状态,D值的正负反映了种群内杂合子的过剩或缺失状态,D值为正表明杂合子过剩,为负表明杂合子缺失[17]。本研究中江鳕群体的D值为0.361 9,表明其杂合子过剩,说明江鳕群体遗传变异程度较大,遗传多样性较高。同时江鳕群体的有效等位基因数Ne为1.833 3,这比报道的硬骨鱼类的多样性的有效等位基因数(Ne=1.16~1.58)[18]的上限略高,可能与本研究中同工酶样本数量不多有关。额尔齐斯河江鳕群体的种质资源库具有较大的变异程度,因此额尔齐斯河江鳕可为后续的人工育种提供良好的亲本。
同时本研究通过对江鳕6种主要组织的同工酶凝胶电泳图谱分析,初步确定了IDHP在肌肉组织中的表达量高且活性稳定,因此异柠檬酸脱氢酶可作为江鳕鉴定的生化遗传标记。
该研究通过对江鳕多种同工酶在不同组织中的差异表达分析,有利于从生化遗传的角度判定江鳕与其他属鱼类亲缘关系的远近,同时也可为江鳕种质标准的建立及良种选育提供理论依据。
猜你喜欢 电泳脱氢酶鱼种 传说中的快速解酒方法有效吗?家庭科学·新健康(2021年5期)2021-06-21古老鱼种重返伊利诺伊州水域疯狂英语·爱英语(2020年12期)2020-12-29脱氢酶活性检测方法及其在环境监测中的应用科学与财富(2020年1期)2020-03-02鱼种优劣鉴别方法农村百事通(2019年2期)2019-03-26喝酒“脸红”和“脸白”,哪个更能喝家庭医药·快乐养生(2017年11期)2017-11-21冬放鱼种技术科学种养(2017年1期)2017-02-16转基因大豆秸秆还田对土壤酶活性的影响江苏农业科学(2016年7期)2016-10-20鲈鱼骨酸溶性和酶溶性胶原的性质比较河北渔业(2015年10期)2015-10-15鱼种深秋管理四要点农家科技(2014年11期)2015-03-11宇通展示新引进的整车电泳设备人民交通(2009年5期)2009-06-01推荐访问:遗传学 生化 遗传推荐文章
- 2018年江西赣州市医疗急救中心招聘编外人员公告:赣州市医疗急救中心
- 升学宴策划活动方案|2018升学宴活动方案
- 小学六一游园活动总结 [小学六一游园活动策划]
- 2018年中国工商银行广东分行暑期实习生招聘岗位、报名时间:2018中国工商银行广西分行春招
- 澳洲留学八大名校排名申请条件_澳洲留学奖学金申请条件及时间
- [调工商档案介绍信范文] 工商档案查询介绍信
- 加拿大亲属移民政策最新更新|加拿大亲属移民条件
- 初一下册语文练习册答案人教版2018 2018人教版语文练习册答案
- 贵州贵阳房价2018 2018年贵州贵阳中医学院第二附属医院招聘方案
- 【2018广东省湛江市赤坎区审计局招聘公告】2018湛江市赤坎区教师招聘